• HIGHLIGHTS
- The SLC2A9, LRRC20, PLK1, and AATK genes can be a novel treatment target for diabetic erectile dysfunction (ED).
- Hippocampal dysfunctions may be related to the alteration of structural plasticity in Parkinson disease animal models, possibly by a decrease in DAergic signaling.
INTRODUCTION
Diabetes mellitus (DM) is a global epidemic that is currently affected 18.2 million people in the Unite State alone [1]. Diabetes was estimated to affect more than 225 million people by 2010, and it is projected to affect 300 million by 2025 worldwide [2-4]. Diabetes involves numerous end-organ complications.
The prevalence of erectile dysfunction (ED) is 3 times higher in men with diabetes than the general population, with up to 3-quarters of diabetic men suffering from it. Men with diabetes also experience an earlier onset of ED [5].
ED is associated with many factors, ranging from psychosocial disorders, nerve injury, smooth muscle, and vascular pathologies [6,7]. Although there have been recent studies detailing the mechanisms of impaired diabetic ED [8], it is likely that the genetic alterations that have been highlighted thus far only account for a limited proportion of the alterations to the erectile tissues that are associated with DM. The severity of ED is also increased in diabetic men. Lastly, existing drug-based ED treatments are less effective for this group [9-11]. Knowing the upregulated or downregulated genes of diabetic ED would allow researchers to inhibit or enhance the expression of such genes by using RNA or gene overexpression.
The streptozotocin (STZ)-induced DM rat is a well-established animal model that recapitulates many aspects of the human urological complications, including ED [12]. Thus, we designed the present study to investigate upregulated or downregulated genes in cavernous tissue from rats with STZ-induced diabetic ED.
MATERIALS AND METHODS
Animals and Induction of Diabetes
We kept Sprague-Dawley male rats weighing 250 g in a temperature-controlled cage. We induced DM in the rats through intraperitoneal injection of 60 mg/kg STZ (Sigma Chemical, Seoul, Korea) in a sterile citrate buffer; we administered an injection of the same concentration of citrate buffer to the control rats. We used an Accu-Chek glucose monitor (Roche Diagnostics, Indianapolis, IN, USA) to verify the blood glucose levels by a tail stick one day following injection. We deemed blood glucose exceeding 300 mg/dL to be indicative of DM [13]. We checked the rats’ body weight and glucose levels weekly throughout the 10-week duration of the research.
Erectile Function Assessment
We selected rats for which ED induction was confirmed among DM rats. According to the minimum number for microarray analysis, 4 rats were selected for each group. We measured intracavernosal pressure (ICP) in response to cavernous nerve stimulation on control rats (n=4) and rats with diabetes at 10 weeks following DM induction (n=4). We first used 5% isoflurane vaporized in 100% oxygen to anesthetize the rats, and then we administered 1.5% isoflurane to maintain anesthesia throughout the examination. We continuously observed the arterial blood pressure by inserting a cannula in the right carotid artery.
At this point, we exposed the penis and mobilized the corpus spongiosum to insert a 25G needle into it. We used polyethylene-50 tubing to connect a needle to a pressure transducer containing heparinized saline. We then surrounded the cavernous nerve with bipolar platinum wire electrodes and administered a one-volt stimulation at 12-Hz frequency with square-wave duration of 1 millisecond for 60 seconds, as in the previously reported study [14,15]. We measured the maximal ICP and mean arterial pressure (MAP) during tumescence. Fluctuations in systemic blood pressure were normalized by determining the maximal ICP-to-MAP ratio.
Every assessment is presented in a flowchart (Fig. 1).
Sampling of Tissue
The penis was quickly dissected free at the crura upon completion of the erectile function assessment. We removed any covering skeletal muscle, then we extracted equal segments of the crura and shaft and placed them into 1.5-mL tubes, at which point they were flash-frozen in liquid nitrogen before being stored at -80°C.
RNA Isolation
We employed a mixture of TRIzol reagent (Invitrogen, Waltham, MA, USA) and RNeasy columns (Qiagen, Hilden, Germany) to conduct total RNA isolation. While following the manufacturer’s guidelines, we used RNAlater-ICE (Ambion, Austin, TX, USA), to slowly thaw the penile tissue that had been kept at -80°C. We then used the TRIzol reagent to accomplish tissue homogenization. We added chloroform and used Phase Lock Gel Heavy 2-mL Eppendorf tubes to conduct centrifugation to allow for better recovery of the aqueous solution phase. We introduced a mixture of one half-volume of 100% ethanol and the recovered aqueous solution and introduced it into a RNeasy mini column. We employed the RNase-Free DNase Kit (Qiagen) to perform on-column DNase digestion, and we used the 2100 bioanalyzer (Agilent Technologies, Santa Clara, CA, USA) to assess the quality of the RNA using RNA 6000 LabChip Kit (PerkinElmer, Waltham, MA, USA). We measured the amount of total RNA and cRNA and absorbance at 260/280 nm using an ultraviolet spectrophotometer. Lastly, we separated the quantified RNA samples into 5-μg aliquots and placed them in storage at -80°C [16].
Hybridization of Microarray and Analysis of Data
We used a Low RNA Input Linear Amplification kit PLUS (Agilent Technologies) for amplification and labeling of the doublestranded cDNA. We used a Gene Expression Hybridization Kit (Agilent Technologies) and Gene Expression Wash Buffer Kit (Agilent Technologies) for hybridization and washing purposes. We used 8 GeneChips (Agilent Rat Gene Expression 4 X 44K Gene Chip) for the 4 controls and the 4 diabetic samples. We conducted image processing using a DNA microarray scanner and Feature Extraction software (Agilent Technologies). Intensity-dependent normalization (locally weighted scatterplot smoothing, LOWESS) was performed, where the ratio was reduced to the residual of the LOWESS fit of the intensity vs. ratio curve. The averages of normalized ratios were calculated by dividing the average of normalized signal channel intensity by the average of normalized control channel intensity. Functional annotation of genes was performed according to Gene Ontology Consortium (http://www.geneontology.org/index.shtml) by GeneSpringGX7.3.1.
Immunohistochemical Staining
We performed immunostaining of the solute carrier family 2 member 9 (SLC2A9), leucine rick repeat containing 20 (LRRC20), polo like kinase 1 (PLK1), and apoptosis-associated tyrosine kinase (AATK) protein on the cavernosum sinusoids. We performed Masson trichrome (Sigma Diagnostic, St. Louis, MO, USA) staining for collagen on 5-μm tissue sections derived from penile shaft. For the detection of apoptosis, we carried out terminal deoxynucleotidyl transferase mediated dUTP nick end labeling (TUNEL) assay with a terminal deoxynucleotidyl transferase (TdT) apoptosis detection kit (Takara BIO Inc., Mountain View, CA, USA) and a DAB Detection IHC Kit (Abcam, Cambridge, UK). We subjected the sections to incubation with 20-μg/mL proteinase K, 2% hydrogen peroxide to quench endogenous peroxidase activity, digoxigenin-conjugated nucleotides and TdT, and antidigoxigenin peroxidase. We used 0.5% diaminobenzidine/0.01% hydrogen peroxide for staining of the sections and hematoxylin for counterstaining. For the negative control, we used the buffer instead of the TdT enzyme. Quantitative analysis of the stained area calculations was performed using Image J version 1.8.0_172 (nih.gov).
Statistical Analysis
For analysis, we entered the raw data into GeneSpring GX 7.0 software (Agilent Technologies). We used this software to produce P-values with which we determined gene detection based on 4 biological replicates from every empirical condition. Meanwhile, we determined the differential expression of genes in both the control and diabetic groups based on the analysis of variance-derived P-values. Fold change was approximated for every gene in the control group. These variables made it possible to subject the array data to statistical filtering. We performed statistical filtering of the array analysis with the R package Linear Models for MicroArray Data (LIMMA) (Accessed: Jun 07, 2021, Bioconductor.org), background correction and normalization of microarray data. We performed Bonferroni correction to adjust the P-values. We carried out t-test to determine whether the groups differed significantly, with data being expressed as mean±standard error and statistical significance denoted by a P-value of less than 0.05.
RESULTS
Glucose Levels in Diabetic Rats
Diabetic rats demonstrated a significantly lower body weight following 10 weeks of SZT injection compared to control rats (288±17 vs. 470±10 g, respectively, P<0.01). Diabetic rats following 10 weeks of SZT injection demonstrated significantly higher blood glucose level than the controls (554±26 vs. 106±2 mg/dL, respectively, P<0.0001), and the group that had been administered STZ exhibited glucose levels of 300 mg/dL or higher throughout the entire research period.
Erectile Function in Diabetic Rats
Ten weeks following SZT injection, diabetic rats showed a reduction in erectile function when the cavernous nerve had been stimulated (Fig. 2), as well as a lower ratio of ICP-to-MAP (0.3537±0.052) compared to the controls (0.86±0.054) (P< 0.01). We therefore concluded that the SZT injections induced diabetic ED in diabetic rat (Fig. 2).
Array Analysis and Measures for Assessing Biological Significance
Statistical filtering of the array analysis showed 9,623 upregulated genes and 2,013 downregulated genes with at least a 2-fold change in expression (P<0.05).
To investigate how biologically significant the 11,636 genes with upregulation or downregulation were for diabetic ED, we adopted a variety of approaches. Many genes are represented by more than one Agilent Rat Gene Expression 4 X 44K Gene Chip and more than one probe set for the same gene was established to have significance in numerous cases. We subjected those genes to a differential expression analyzed via GeneSpring GX 7.0 (Agilent and DAVID bioinformatics tools, National Institute of Allergy and Infectious diseases) and Gene Ontology and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis. This facilitated gene classification based on biological process, molecular function, and subcellular location. Table 1 and Fig. 3 show the KEGG pathways for the differential expressed genes.
According to the GO analysis, there was a significant enrichment of 2,013 downregulated genes in the GO categories including the following: cell cycle 16 (0.8%), extracellular matrix (ECM)-receptor interaction 10 (0.5%) conferring tensile strength, and glycosylphosphatidylinositol (GPI)-anchor biosynthesis 5 (0.2%). There was also an enrichment of 9,623 upregulated genes in the GO categories, including apoptosis 32 (0.3%), arginine and praline metabolism 25 (0.3%), gap junction 54 (0.6%), p53 signaling pathway 23 (0.3%), regulation of actin cytoskeleton 66 (0.7%), transforming growth factor (TGF)-β signaling pathway 34 (0.4%), tight junction 53 (0.6%), vascular smooth muscle contraction 54 (0.6%), and vascular endothelial growth factor (VEGF) 29 (0.3%) signaling pathway.
From 41,105 genes, statistical filtering of the array analysis showed 9,623 upregulated genes and 2,013 downregulated genes with at least 2-fold changes in expression (P<0.05). According to R package LIMMA, the number of 2-fold changes with all 4 DM penis samples was found to be 1,099. According to the ttest, the number of all 2-fold changes was 1000 (P<0.05). With Bonferroni correction (P<4.55×10-5), the number of all 2-fold changes was 4: SLC2A9, LRRC20, PLK1, and AATK (Fig. 4).
Immunohistochemical Staining
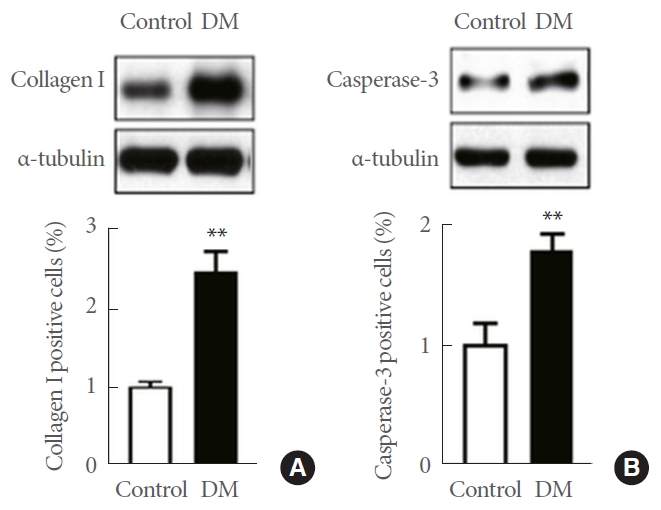
DM group showed increased expression of collagen 1of penile cavernosum than control group. DM group showed increased expression of caspase-3 of penile cavernosum than control group (Fig. 5).
To determine where in the cavernosal tissue the AATK, SLC2A9, and PLK1 proteins are located, we conducted immunofluorescence staining. As shown in Fig. 6, the cavernosal sinusoids or smooth muscle were lined with tissue layers containing bright AATK, SLC2A9, and PLK1 antibody labeling (green; Alexa 488). It therefore appears that AATK, SLC2A9, and PLK1 are labeling the endothelium and smooth muscle layers of the cavernosal sinusoids. In the diabetic group, cavernosal AATK area increased (Fig. 6D, G), SLC2A9 (Fig. 6E, G), and PLK1 area decreased (Fig. 6F, G), compared to control group (Fig. 6A-C).
DISCUSSION
It is difficult to determine the biological relation of modified genes. GO analysis and National Center for Biotechnology Information (NCBI) literature review were among the approaches we used in the present work to interpret the results obtained from a biological perspective. GO categories (e.g., cell cycle, ECM, and GPI-anchor biosynthesis) exhibited a marked proliferation of downregulated genes. By contrast, upregulated genes proliferated in GO categories, including apoptosis, arginine and praline metabolism, gap junction, p53, regulation of actin cytoskeleton, TGF-β, tight junction, vascular smooth muscle contraction, and VEGF.
With R package LIMMA, we performed background correction and normalization of microarray data. The number of 2-fold changes with all 4 DM penis samples was 1,099. Gene expression data obtained by microarray or RNA-sequencing technologies can be analyzed via the LIMMA package [17], which allows for simultaneous comparisons between multiple RNA targets. Further, the data derived from any technology of quantitative gene expression (e.g., microarrays, RNA-sequencing, quantitative polymerase chain reaction) can be subjected to the linear model and differential expression functions [18].
Neuronal cell proliferation, differentiation, synaptic transmission, and plasticity, along with survival and death, are among the neuronal processes regulated through the involvement of numerous protein kinases. The AATK, protein kinase, is primarily expressed in the nervous system and contributes to cell apoptosis and neurite growth. Diabetic ED involves various neural mechanisms [19,20]. As AATK could involve the penile cavernous nervous system, it is reasonable that diabetic ED may develop as a result.
The downregulation of VEGF may contribute to diabetic penile vasculopathy [21]. In the present study, the gene of SLC2A9 (GLUT 9) was down regulated in diabetic ED (Fig. 4) and the SLC2A9 protein was demonstrated to be localized to the sinusoids of the cavernosum. Its area decreased (Fig. 6). In this work, diabetes-related ED was associated with alterations in the SLC2A9 gene. Evidence was provided that the cavernosal sinusoids contained the SLC2A9 protein (Fig. 6). Encoded by the SLC2A9 gene, the SLC2A9 protein belongs to the solute carrier family 2 and mediates the glucose transporter member 9 [22]; this family is therefore critical for the maintenance of glucose homeostasis. Further, SLC2A9 is a transporter of uric acid, and a correlation has been established between genetic variants of the transporter and a greater predisposition to hyperuricemia, which is related to the development of endothelial dysfunction [23-25].
In the present study, the gene of PLK1 was downregulated in diabetic ED (Fig. 4). The PLK1 protein was demonstrated to be localized to the sinusoids of the cavernosum. Its area decreased (Fig. 6). This protein contributes to the regulation of mitosis and plays a central role in the processes of centrosome maturation, kinetochore-microtubule adhesion, development of bipolar spindles, and cytokinesis [26]. The loss of PLK1 expression can induce pro-apoptotic pathways and inhibit growth. As PLK1 could involve adaptive pancreatic beta cell proliferation and blood glucose levels, it is reasonable that diabetic ED may develop as a result [27].
Gene of LRRC20 (leucine rich repeat containing 20), a member of the leucine rich repeat superfamily of proteins, were downregulated in diabetic ED (Fig. 4). The LRRC20 is a transmembrane protein that is encoded by the LRRC20 gene. The function of LRRC20 is not well known, and it is speculated to involve cell proliferation or differentiation [28].
Further, the Agilent Rat Gene Expression 4 X 44K Gene Chips comprise probe sets denoting genes and transcripts, including most full-length, well-annotated rat genes. However, they do not include probe sets for every rat gene. Therefore, the present analysis is not comprehensive. Moreover, gene expression was assessed at one point in time, which means that we might have overlooked important modifications occurring at an earlier or later point in ED evolution. Thus, ED-related genes might be more effectively identified through an inclusive analysis of temporal alterations according to expression patterns over time.
Determining the biological role and impact of altered genes is challenging microarray is a method to explore the gene expression situation comprehensively and is helpful in identifying changes in genes, such as increase or decrease in specific mRNA [29]. In addition, it is easy to find genetic differences that are expressed differently under different conditions. GO analysis and NCBI literature review were among the approaches used to interpret the results obtained from a biological point of view. Fold change methods are measures that describe the change in a quantity between an original measurement and a subsequent measurement, defined as the ratio between 2 quantities [29]. This method is a practical approach for selecting differentially expressed genes from microarray data and has the advantage of being easy to interpret. However, the data is biased, and values that are too asymmetric with respect to 1 may be obtained. Also, the value of fold change may be disproportionately affected by measurement noise.
Our study has some limitations. Despite the benefits associated with using microarrays for expression analysis, the method also has certain drawbacks. Various types of cells are present in the erectile tissues in the extracted cavernous samples, which could affect the observed differential gene expression. Further, this work was limited to the formulation of a hypothesis, as it did not go beyond confirming that the gene modifications and diabetic ED were correlated. Therefore, additional empirical research must be conducted to identify which genes with differential expression mediate ED, which genes are ED indicators, and which genes are unrelated to ED. The present work should be followed by a knock down of selected genes with an in vitro cell culture study and a knockout of selected genes with an in vivo study for the confirmation of the functions of the selected genes. Moreover, cohesive pathway analysis has not been performed in this study. Due to the limited number of DM induced rats, we could not expect the accurate pathway using cohesive analysis.
This study used the SZT-induced DM model, which is a type I DM model. Type I DM is caused by the destruction of pancreatic beta cells, leading to insulin deficiency. Autoimmune mechanisms cause most cases of type I DM. However, it is not a common form of diabetes. In type 2 DM, insulin resistance increases, insulin sensitivity decreases, and blood sugar rises. Hereditary obese diabetic mice, mainly including leptin receptor knockouts, are widely used in animal studies. However, type 2 DM is a disease caused by a combination of accumulated polygenic traits and interactions with environmental stressors. As a complex and heterogeneous disease caused by a combination of genetic and lifestyle factors, no single model is most suitable for all type 2 DM models. STZ gives nonspecific organ toxicities [30]. Although type I DM model used in this study does not represent general DM, the STZ-induced DM model has already been used in several diabetic mouse experiments [30].
In another study on STZ-induced ED in diabetic rats, diabetic rats showed lower VEGF immunoexpression and higher TGF-β immunoexpression than normal controls [31]. This result is contrary to our study. There are many levels of regulation between transcription and translation. An increase in mRNA does not always mean an increase in protein levels [32]. This study did not reveal how these genes work or possible mechanisms. However, the altered gene expression we identified could be considered a new therapeutic target and a new target for the treatment of diabetic ED.