Liberation of Serotonin Is Not Unaffected by Acetylcholine in Rat Hippocampus
Article information

Abstract
Purpose
Raised cerebral titers of acetylcholine have notable links with storage symptomatology related to lower urinary tract symptoms. The hippocampus contributes to the normal control of continence in the majority of instances (circuit 3). Owing to synaptic connections with other nerve cells, acetylcholine affects the micturition pathway via the liberation of additional cerebral neurotransmitters. Despite the fact that cerebral serotonin is a key inhibitor of reflex bladder muscle contractions, the influence of acetylcholine on its liberation is poorly delineated. The current research was conducted in order to explore the role of acetylcholine in serotonin liberation from sections of rat hippocampus in order to improve the comprehension of the relationship between cholinergic and serotonergic neurons.
Methods
Hippocampal sections from 6 mature male Sprague-Dawley rats were equilibrated over a 30-minute period in standard incubation medium so as to facilitate [3H]5-hydroxytryptamine (5-HT) uptake. The cerebral neurotransmitter, acetylcholine, was applied to the sections. Aliquots of drained medium solution were utilized in order to quantify the radioactivity associated with [3H]5-HT liberation; any alterations in this parameter were noted.
Results
When judged against the controls, [3H]5-HT liberation from the hippocampal sections remained unaltered following the administration of acetylcholine, implying that this agent has no inhibitory action on this process.
Conclusions
Serotonin liberation from murine hippocampal sections is unaffected by acetylcholine. It is postulated that the bladder micturition reflex responds to acetylcholine through its immediate cholinergic activity rather than by its influence on serotonin release. These pathways are a promising target for the design of de novo therapeutic agents.
• HIGHLIGHTS
- Raised cerebral titres of acetylcholine have notable links with storage symptomatology related to lower urinary tract symptoms.
- Serotonin liberation from murine hippocampal sections is unaffected by acetylcholine.
- It is postulated that the bladder micturition reflex responds to acetylcholine through its immediate cholinergic activity rather than by its influence on serotonin release.
INTRODUCTION
Alzheimer disease (AD) is the most frequent underlying etiology of dementia in older individuals. The symptomatology of this condition has been postulated to stem from degradation of cholinergic nerve cells and diminished cholinergic neurotransmitter release [1]. A cholinesterase inhibitor, donepezil has been demonstrated to elevate acetylcholine concentrations and therefore heighten cerebral function in patients suffering from AD [2]. Nevertheless, elevated titres of acetylcholine are notably linked to lower urinary tract symptoms (LUTS) storage issues. The possibility of deteriorating urinary incontinence is a key factor when initiating cholinesterase therapy in this form of dementia [3].
As a neurotransmitter, acetylcholine displays both suppressive and stimulatory activity, according to the form of receptor that is triggered and its site within the cerebrum [4,5]. Owing to synaptic connections with additional neurons, acetylcholine moderates the micturition pathway through the liberation of additional cerebral neurotransmitters. Acquiring information relating to these neurochemical cues within the central pathways that govern bladder control is essential in order to comprehend the way in which these circuits work and how they can be influenced for the therapy of the storage symptomatology of LUTS. These pathways appear to have potential for the design of de novo pharmaceutical agents.
The configuration of the hippocampus contributes to continence in healthy people for the majority of the time (circuit 3) [5-7]. Both serotonergic and cholinergic neurons pass into the hippocampus from the raphe or medial septal nuclei, respectively. Numerous studies have been performed so as to evaluate their internal and external links [8-12]. Nevertheless, the acetylcholine network involved in the cerebral moderation of serotonin on the micturition pathway has not been fully delineated. The current work was conducted in order to explore the role of acetylcholine in the modification of serotonin liberation from sections of murine hippocampus.
MATERIALS AND METHODS
Preparation of Hippocampal Slices
All study protocols were performed in keeping with the National Institute of Health Guide for the Care and Use of Laboratory Animals (2001); they were sanctioned by the facility’s Institutional Animal Care and Use Committee.
The rodents utilized in this study were adult male SpragueDawley rats, weighing between 150 and 200 g. They were accommodated to the laboratory surroundings for a minimum of a week. The rodents underwent decapitation; the cerebra were swiftly extracted and then carefully separated at the midline into 2 hemispheres. Isolation of both hippocampi was performed and each hippocampus was sectioned to generate 400- µm slices along the transverse plane using a Mcilwain mechanical tissue chopper (The Mickle Laboratory Engineering Co., Gomshall, Surrey, UK).
These processes were all conducted on ice; the tissues were repetitively rinsed with routine incubation medium at an ice cold temperature of between 2°C and 4°C. The medium comprised NaCl 124mM, KCl 4mM, CaCl2 2mM, MgSO4 1mM, KH2PO4 1.25mM, NaHCO3 25mM, glucose 10mM, and was modified to a pH within the range 7.3–7.5 with 95% O2/5% CO2. 12.5μM nialamide (Sigma Chemical Co., St. Louis, MO, USA) was admixed in order to inhibit the metabolic breakdown of 5-hydroxytryptamine (5-HT).
Experimental Protocol
Equilibration of 6 sections slices of murine hippocampus was conducted over 30 minutes in the standard incubation medium heated to 37°C and saturated with 95% O2/5% CO2. After this time interval, the first medium was substituted for a de novo solution which included [3H]5-HT (0.1μM, 74μCi; Amersham International plc., Buckinghamshire, UK) in order to achieve serotonin uptake.
Following a 20-minute incubation period, the sections were thrice cleansed with the standard medium. Each section was randomly assigned to a glass tube which held a further 3 mL of incubation medium. 10μM zimelidine (Research Biochemical International, Natick, MA, USA) was included in the medium in order to suppress reuptake during the study. This fluid was then collected and a de novo aliquot of standard incubation medium was added to the tubed specimen; this process was repeated every 10 minutes for 100 minutes. The radioactivity of the liberated 5-HT was quantified from the drained solution. 50 minutes after the beginning of the observation period, the pharmaceutical agents under investigation were applied, one at the sixth 10-minute period and the other at the seventh. To determine whether the effect of the neurotransmitters was influenced by interneuron, delivery of tetrodotoxin occurred over the whole 100-minute time interval.
Radioactivity Count of [3H]5-HT
Nine-mililiter scintillation cocktail (Ready Safe, Beckman Instruments Inc., Fullerton, CA, USA) was admixed with a single ml of the medium collected from the ampoule at each 10-minute period. In order to quantify the remaining [3H]5‐HT radioactivity in the sections of hippocampus, 1 mL of tissue solubiliser was employed (Soluene, Pachard Instrument Co., Inc., Downers Grove, IL, USA). This was incubated with the tissues for 120 minutes at a temperature of 37°C. This was then neutralised with 70-µL glacial acetic acid (Merk & Co., Inc., Kenilworth, NJ, USA). One hundred microliter of the entirely solubilised specimen was then combined with a 9-mL volume of liquid scintillation cocktail prior to radioactivity quantification.
Drugs Used
Acetylcholine, tetrodotoxin, and nialamide were obtained from Sigma Chemical Co. (St. Louis, MO, USA). [3H]5‐hydroxytryptamine and zimelidine HCl were acquired from Amersham International plc. (Buckinghamshire, England) and Research Biochemicals International (Natick, MA, USA), respectively. The tissue solubiliser, Soluen 350 m, and the liquid scintillation cocktail, Ready Safe, were sourced from the Packard Instrument Company Inc. (Downers Grove, IL, USA) and Beckman Instruments Inc. (Downers Grove, IL, USA), respectively.
Data Analysis
The results are all stated in terms of fractional release (FR), which was calculated according to the following equation:
where cpm (medium) FR represents the radioactivity (counts per min, cpm) of [3H]5-HT liberated into the media collected during a specific 10-minute fraction, Cpm (medium) total indicates the sum of the radioactivity of [3H]5-HT discharged into the media collected at each 10-minute interval from the noted time fraction to the observation period’s conclusion, and Cpm (tissue) is defined as the residual radioactivity of [3H]5-HT on the hippocampal sections at the end of the 100-minute observation interval.
Analysis of variance was employed for the statistical analysis relating to comparation of the different cohorts; Scheffe test was subsequently performed. A P-value of 0.05 was deemed significant.
RESULTS
Release of [3H]5‐HT in Control Group and Tetrodotoxin Preadministered Group Utilizing Standard Media
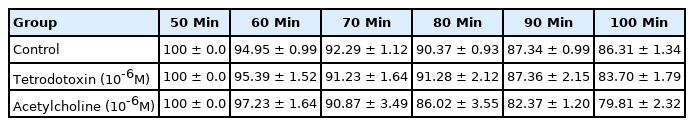
Spontaneous liberation of 5-HT from the sections of campus was demonstrated during the entire experiment (Table 1); the rate diminished quickly over the initial 40 minutes but was then followed by a constant release for the remainder of the 100- minute time interval. 5-HT was unaffected by therapy with tetrodotoxin (10-6M) (Fig. 1). The pharmaceutical agents under investigation were given for a 20-minute period commencing at the 60- and 70-minute time points, respectively. The liberation of 5-HT at the fifth 10-minute interval was taken as a reference point; any alterations in 5-HT secretion were reported as a percentage of this value.
Effect of various neurotransmitters on the % release of 5-hydroxytryptamine

Spontaneous release of [3H]5-hydroxytryptamine.
Change of [3H]5-HT Release by Acetylcholine
In order to investigate the action of acetylcholine on 5-HT liberation from the sections of hippocampus, the incubation medium was exposed to this agent for 20 minutes. At the 60-minute juncture, the control cohort exhibited a [3H]5-HT release of 94.9%±1.0%; at the 70-minute time point, the equivalent figure was 92.3%±1.1% (Table 1). The acetylcholine (10-5M) had no impact on the liberation of [3H]5-HT by the hippocampal specimens’ (Fig. 2).

Effect of acetylcholine(10-5M) on the % change of 5-hydroxytryptamine (5-HT) release.
DISCUSSION
Immediate synaptic feeds to the sacral preganglionic nerve cells or sacral dorsal commissure area are offered by nerve cells within the pontine micturition centre. These are believed to be key in moderating either a triggering or suppressive effect on urine release or continence [13,14]. Nevertheless, the stimulation of the subcortical network, which encompasses the periaqueductal gray and parahippocampal cortex, arises in the absence of sensation (circuit 3) in the majority of normal scenarios in which the bladder becomes full steadily. Given that the parahippocampal cortex lies in proximity to the amygdala, this circuit is predominantly related to the safety and emotional facets of releasing urine, possibly offering output to the nuclei in the brainstem through the hypothesised safe cue of continence arising from the hypothalamus [5-7].
A range of neurotransmitters is thought to be involved in the cerebral governance of the lower urinary tract. The key enabling transmitter appears to be glutamate; this acts within the spinal and supraspinal reflex mechanisms that govern the bladder and the external urethral sphincter [5,13-15]. The principal neurotransmitter that inhibits central governance is 5-HT, also known as serotonin. There is a heavily populated serotonergic nerve network within the spinal reflex circuits associated with the action of bladder emptying. Stimulation of the central serotonergic system can inhibit this process by suppressing the excitatory parasympathetic influence on the bladder; 5-HT produces an extended triggering time for the thoracic sympathetic preganglionic nerve cells. In cats, raphe nuclei activation suppresses reflex bladder function [5,16-18].
Within the cerebrum, acetylcholine may act as an inhibitor or stimulator [5,8]. An inhibitory influence on the spinal cord’s micturition reflex is mediated by muscarinic acetylcholine receptors, whereas an excitatory impact on this reflex in murines is seen via nicotinic receptors. A single neurotransmitter has the capacity to moderate additional cerebral neurotransmitter liberation owing to synaptic connections with neighboring nerve cells. Thus, the cholinergic system impacts the micturition process via serotonin liberation together with cerebral cholinergic pathways. The hippocampus receives serotonergic and cholinergic neurons from the raphe nucleus and medial septal nucleus, respectively [4,15,19-21]. They then project across the majority of the hippocampus, a feature which is particularly evident in the dentate gyrus [5,8-11]. Many cerebral nerve cell cohorts contribute to bladder, urethra, and urethral sphincter governance, encompassing the raphe nuclei in the medulla, which include serotonergic nerve cells [22-24].
In this experiment, no variation in 5-HT liberation was seen in either the control cohort or the slices to which acetylcholine was administered with respect to a concentration of the latter of up to 10‐5M. The serotonergic nerve cells were uninfluenced by their cholinergic counterparts. Serotonin liberation was not suppressed by a higher concentration of acetylcholine. It is postulated that although acetylcholine impacts the bladder micturition reflex, this action is not mediated via its influence on serotonin liberation but through the immediate effect of acetylcholine on muscarinic receptors.
In conclusion, liberation of 5-HT from murine hippocampal sections was unaffected by acetylcholine. It is proposed that acetylcholine’s role in the bladder micturition reflex arises through its immediate action on cholinergic muscarinic receptors rather than via any influence on serotonin liberation. These pathways are promising targets for the development of de novo therapeutic agents.
Notes
Fund/Grant Support
This research was supported by grant from Soonchunhyang University Research Fund and the National Research Foundation of Korea (NRF) funded by the Ministry of Education, Science and Technology (2021R1A2C1004163).
Research Ethics
This research was approved by the Institutional Animal Care and Use Committee of Soonchunhyang University Seoul Hospital (IRB No. 2019-4). The authors carried out the experimental protocols, in line with the National Institute of Health Guide for the Care and Use of Laboratory Animals.
Conflict of Interest
No potential conflict of interest relevant to this article was reported.
AUTHOR CONTRIBUTION STATEMENT
·Conceptualization: YSS
·Data curation: JHK, YSA
·Formal analysis: YSA, YSS
·Funding acquisition: YSS
·Methodology: JHK, YSA
·Project administration: YSS
·Visualization: YSA
·Writing-original draft: JHK, YSS
·Writing-review & editing: YSA, YSS