INTRODUCTION
Pioglitazone is one of thiazolidinediones (TZDs), which is currently the only oral drugs with a confirmed antihyperglycemic impact in type 2 diabetes mellitus (T2DM) patients. TZDs is well-known agonists of peroxisome proliferator-activated receptor (PPAR) α and γ, TZDs are known to positively influence insulin sensitivity and β-cell function and potentially alter lipid profiles. Pioglitazone is a synthetic ligand of PPARγ and is used as a therapeutic treatment for patients with T2DM. PPARγ, a ligand-activated transcription factor, is expressed in white and brown adipose tissues as well as in the urinary bladder [1]. Pioglitazone has also been shown to reduce macrophage infiltration through the activation of PPARγ and induction of apoptotic cell death [2], which can lead to decreased numbers of macrophages in adipose tissues [3]. To examine the impact of PPARγ on bladder cell function, several studies compared the PPARγ levels in normal bladder and bladder cancer (BC) mucosal samples. They found that cell migration and invasion rates are correlated with PPARγ expression in various BC cell lines. Recently, in vitro effects of pioglitazone on normal urothelial epithelium (NUTE) cells and BC (J82) cells were compared [4]. In this paper, pioglitazone inhibited the proliferation and induced apoptosis of NUTE cells, but not J82 cells, suggesting the different effects of pioglitazone on normal bladder cells vs BC cells. In addition, prior studies showed that a primary proangiogenic factor, vascular endothelial growth factor A, is positively regulated by PPARγ [5]. PPARγ is involved in proliferation and survival through Wnt/β-catenin, IGF-I/PI3K/AKT/mTOR, and mitogen activated protein kinase (MAPK) pathways [6]. Furthermore, PPARγ can regulate the inflammatory response of macrophages through Toll-like receptor 4/nuclear factor-κB (TLR4/NF-κB) signaling [7].
There have been contradictory reports on diabetes mellitus (DM) and elevated risk of BC [8]. Studies have demonstrated that pioglitazone can have adverse effects that exaggerate the risk of BC. DM patients, particularly males, on long-term, high doses of pioglitazone, are at a higher risk of developing BC [9]. The KPNC (Kaiser Permanente Northern California) study determined that inflated dose and length of pioglitazone treatment correlated to increased rates of BC [5]. Currently, the U.S. Food and Drug Administration allows the use of pioglitazone in patients with special caution to those with a prior history or active BC [10]. A potential reason for this observation could be that PPARγ signaling in BC cells may provide a tumor micro-environment that allows for de novo lipogenesis of lipids that can be utilized in increasing tumor mass and energy usage [11]. However, other studies have shown that TZDs posed no risk on survival in BC patients who underwent radical cystectomy [12]. A recent meta-analysis found no difference in BC incidence among users of pioglitazone and nonusers [13]. These inconsistencies between studies have led to questions in the legitimacy of studies and the actual dangers of continued pioglitazone use. Due to the inherent difficulty of accurately assessing the link between TZDs and BC, the association between the two is still in controversy [14].
The goal of our study was to determine whether TZDs have any negative or positive effects on the healthy bladder epithelium. Since pathophysiological changes require the actions of a series of key proteins, global proteomic analysis of protein alterations using state-of-the-art mass spectrometry is ideal for acquiring unbiased biological information regarding molecular mechanisms. Using such a method, this study aimed to understand the mechanistic meanings of proteome perturbations in the normal bladder due to TZD treatment. We sought to examine the effects of pioglitazone specifically, being that is the most widely used TZD. Our global quantitative proteomic analyses found that pioglitazone alters the proteome and biological networks in normal human bladder cells without tumorigenic effects.
MATERIALS AND METHODS
Cell Culture
We used human telomerase reverse transcriptase-immortalized human urothelial cell line (TRT-HU1) for this study [15]. Cell lines that were initially established, frozen, and under a passage number of 10 were used in this study. Culture media was changed after one day of subculturing and cells were passed again when there was 70%–80% confluence. All cells were negative for mycoplasma contamination. For maintaining TRTHU1, we used Dulbecco’s modified Eagle’s medium supplemented with 10% fetal bovine serum (Invitrogen, Waltham, MA, USA), 1% penicillin/streptomycin, and 1% L-glutamine (Sigma-Aldrich, St. Louis, MO, USA) under a humidified atmosphere of 5% CO2 at 37°C.
Antibodies and Reagents
Commercial antibodies were obtained from the following sources: actin gamma 2 (ACTG2) (ab189385), myosin heavy chain 3 (MYH3) (ab124205), oxidative phosphorylation (OXPHOS) (ab110413) from Abcam; phospho-NF-κB (3033), phospho-HER2/ErbB2 (2247), phospho-p21 activated kinase 1 (PAK1) (2601), phospho-glycogen synthase kinase-3β (GSK-3β) (9336), Snail (3879), N-cadherin (13116), Slug (9585), β-catenin (8480), E-cadherin (14472), Tight Junction Antibody Kit (8683) from Cell Signaling Technology, and β-actin (A1978) from Sigma. Horseradish peroxidase (HRP)-conjugated secondary antibodies (7074, 7076) were purchased from Cell Signaling Technologies. Most of other chemical reagents were procured from Sigma Chemical Corp.
Quantitative Proteomics
For this study, we performed the tandem mass tagging (TMT)-based analysis as we described in previous papers [15]. In brief, cellular proteins from 25μM pioglitazone-treated and control TRT-HU1 cells were extracted using a 4% sodium dodecyl sulfate-containing buffer. The protein concentration was measured using the Pierce 660nm Assay Kit (Thermo Fisher Scientific, Waltham, MA, USA). The kit was carried out using 60 μg of protein from each sample. Samples were digested with trypsin via filter-aided sample preparation and labeled with TMT6plex reagents in parallel. Then, the peptides were reconstituted and desalted via C18 spin columns (Thermo Fisher Scientific). High-pH reversed-phase liquid chromatography using an Ultimate 3000 XRS System (Thermo Fisher Scientific) was used to separate samples. Peptides with >30% precursor ion interference were excluded to minimize inaccurate quantifications caused by precursor ion interference and Proteome Discoverer was used.
Identification of Differentially Expressed Proteins
Proteins that have more than 3 nonredundant peptides in each sample were selected for further analysis using a one sample t-test using the log2 fold changes to compute the significance. False discovery rates (FDRs) of each protein for the 1 sample t-test were then calculated using Storey’s correction method. Differentially expressed proteins (DEPs) were identified based on FDR<0.05 and absolute log2 fold change ≥0.58 (1.5-fold). DAVID ver. 6.8 was used for functional enrichment analysis of DEPs. Significantly enriched cellular processes were selected (P<0.05). Functional classification analysis was performed using PANTHER ver. 11.
Master Regulator Analysis
To identify master regulators, we discerned each DEP and counted their individual partners. The protein-protein interaction information was based on the following 6 interactome databases: the Biological General Repository for Interaction Datasets (BioGRID), the Database of Interacting Proteins (DIP), High confidence protein-protein interactions (HitPredict), the IntAct molecular interaction database (IntAct), the Molecular INTeraction database (MINT), and the STRING database. Fisher exact test was used to calculate the enrichment P-values using. Finally, we selected proteins with P-values lower than 0.01 and prioritized the DEPs based on the number of interaction partners.
Western Blot Analysis
For western blot analysis we used whole-cell lysates. All primary antibodies incubations were done at 4°C overnight. HRP-conjugated secondary antibody incubations were done at room temperature for 1 hour. All western blot experiments were repeated at least 3 times for each antibody and analyzed from different lysates using standard procedures.
Quantification of Reactive Oxygen Species
We measured intracellular reactive oxygen species (ROS) levels by staining with 2′, 7′-dichlorofluorescin diacetate (DCF-DA) and sorting with a flow cytometer (FACSCalibur, Becton-Dickinson, Franklin Lakes, NJ, USA). DCF fluorescence intensity was measured with maximum excitation and emission spectra of 495 nm and 529 nm, respectively.
Cell Proliferation Assay
To determine cell growth during exposure to pioglitazone, TRT-HU1 were seeded in 6-well plates (5×104 cells/well). After treatment with varying doses of pioglitazone (0, 25, 50, or 100μM) (E6910, Sigma-Aldrich) or vehicle, cell numbers were evaluated after 24, 48, and 72 hours. For crystal violet staining, cells were stained with 0.05% solution of crystal violet. After incubation at room temperature for 15 minutes, the wells were washed thoroughly with phosphate buffered saline and fixed with 4% paraformaldehyde at room temperature for additional 5 minutes. For quantitative analysis, 10% acetic acid solution was used to dissolve the stained cells and absorbance at 570–590 nm were measured. All the data are representative of 3 independent trials.
RESULTS
Pioglitazone Perturbs the Proteome of Normal Bladder Epithelial Cells
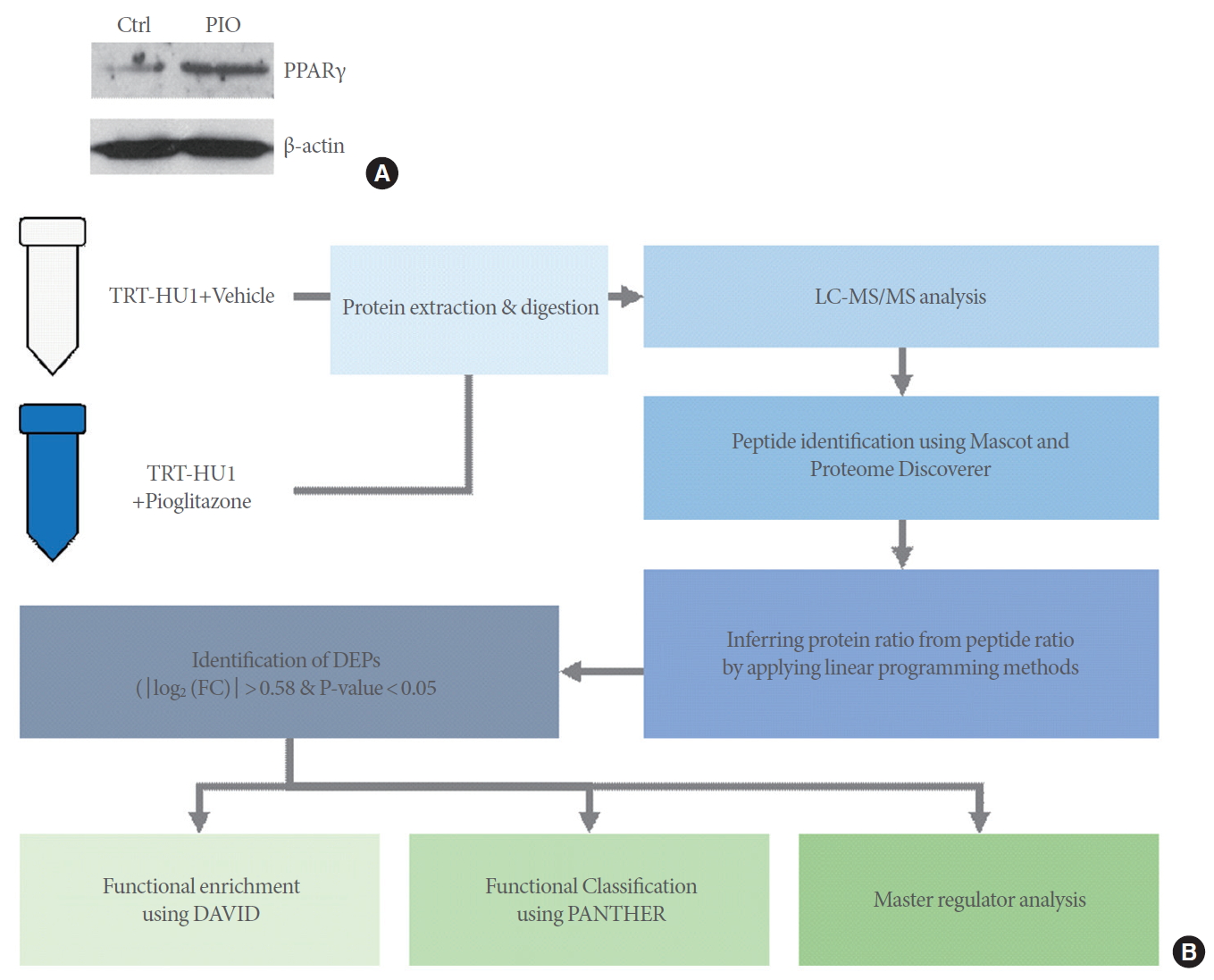
Labeled LC-MS/MS proteomic analysis was conducted to identify the global proteomes of TRT-HU1 bladder cells treated with or without 25μM of pioglitazone. We found that pioglitazone treatment increased PPARγ protein expression (Fig. 1A). Our proteomics analysis workflow is summarized in Fig. 1B.
We detected 5,769 proteins in total, including proteins from the cell part (n=2,375), organelle (n=1,663), macromolecular complex (n=1,060), membrane (n=643), extracellular region (n=88), cell junction (n=61), synapse (n=43), and extracellular matrix (n=29) (Fig. 2A). To identify DEPs between the control versus pioglitazone groups, we performed a statistical hypothesis test using an empirical null model (see Methods). Among the 5,769 proteins, 124 DEPs (including 95 upregulated and 29 downregulated proteins) were found to have absolute log2 foldchanges ≥0.58 and P-values <0.05. Approximately 70% of the DEPs (n=95) showed increased expression due to pioglitazone treatment (Fig. 2B).
Volcano plot displays DEPs between control and pioglitazone groups. The top 5 most significantly up- or downregulated proteins were highlighted with their official symbol (Fig. 2C). HIST1H2BK (histone cluster 1 H2B family member K), ST13P5 (ST13, Hsp70 interacting protein pseudogene 5), and WASH2P (WAS protein family homolog 2 pseudogene) were the most significantly downregulated by pioglitazone treatment. ATPAF1 (ATP synthase mitochondrial F1 complex assembly factor 1), SNX17 (sorting nexin 17), LZTS2 (leucine zipper tumor suppressor 2), and UTP14C (UTP14C, small subunit processome component) were the most upregulated in response to pioglitazone. Supplementary Table 1 shows the list of DEPs that were altered in response to pioglitazone along with the transcriptional targets of PPARγ and test statistics. This data suggest that pioglitazone may induce transcriptional programming through PPARγ, which contributes to increased protein expression.
Functional Enrichment Analysis Reveals That Pioglitazone Modulates Key Biological Processes
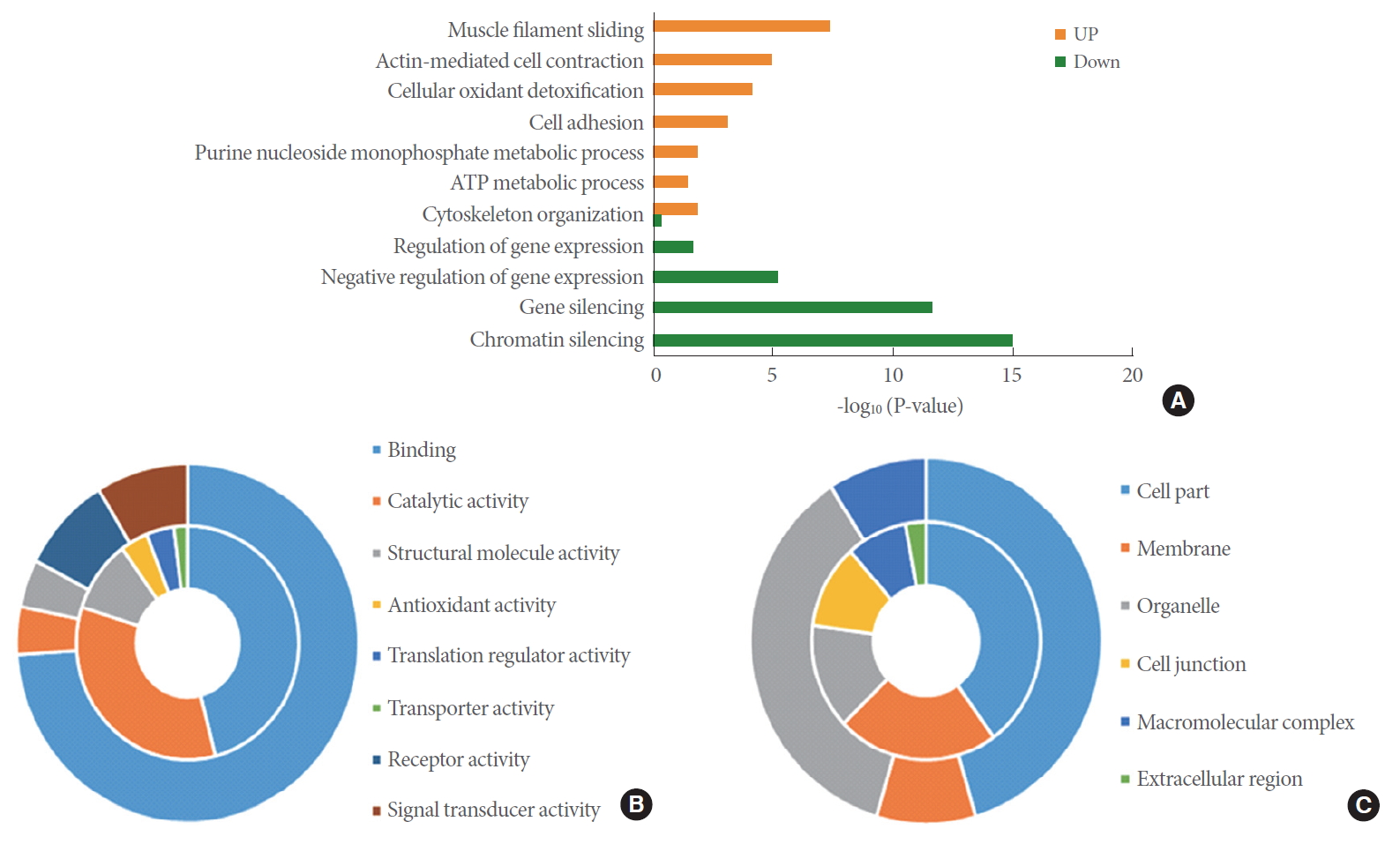
To check the function of proteins perturbed by pioglitazone, we performed functional enrichment and classification using DAVID and PANTHER, respectively. Muscle filament sliding, actin-mediated cell contraction, cellular oxidant detoxification, cell adhesion, purine nucleoside monophosphate metabolic process, ATP metabolic process, and cytoskeleton organization were enriched by upregulated DEPs. However, chromatin silencing, gene silencing, and regulation of gene expression were enriched by downregulated DEPs (Fig. 3A). We listed the DEPs belonging to actin-mediated cell contraction, cellular oxidant detoxification, cell adhesion, purine nucleoside monophosphate metabolic process, and chromatin silencing (Table 1).
Interestingly, the classified molecular functions of the DEPs include binding, catalytic activity, structural molecule activity, antioxidant activity, translation regulator activity, transporter activity, receptor activity, and signal transducer activity (Fig. 3B). Binding function was the largest portion of both the upregulated and downregulated DEPs. Catalytic, antioxidant, translation regulator, and transporter activity were the major molecular functions in the upregulated DEPs. However, receptor and signal transducer activity were the major functions in downregulated DEPs. The classified cellular compartments of the DEPs were cell part, membrane, organelle, cell junction, macromolecular complex, and extracellular region (Fig. 3C). Membrane, cell junction, and extracellular region were the major cellular compartments in upregulated DEPs. Organelle and macromolecular complex are major cellular compartments in the outer circle.
Identification of Protein Interaction Networks Regulated by Pioglitazone Treatment
Our present study revealed a large set of proteins (n=5,769) with relevant levels of abundance in normal bladder epithelial cells. Their functions are related to various cellular processes, including actin regulation and chromatin remodeling (Fig. 3A). Pioglitazone perturbs and modulates these cellular processes by affecting protein-protein interactions. We therefore performed master regulator analysis using all the proteins detected in our global proteomic analysis to identify key proteins that can be major nodes in the protein-protein interaction networks. To select key proteins, we first identified proteins with a large number of interactions from those detected. We then determined the top 5 proteins with the largest number of interactions and the most significant P-values (Fig. 4A, Supplementary Table 2).
To reconstruct the network model, top 3 proteins were then used. Of note, B lymphoma Mo-MLV insertion region 1 homolog (BMI1), Histone H4A (HIST1H4A), and superoxide dismutase 1 (SOD1) were found to be the top 3 regulators, with P-values <0.01 and large target counts. The network model describes the interactions among the top 3 regulators and their interactors with significant differential expression when treated with pioglitazone. Pioglitazone treatment downregulated expression of BMI1, which was found to interact mainly with histone proteins, such as Histone H2 (HIST2A), Histone H2A type 3-A (H2AA3), Histone H2A type 2-C (HIST2H2AC), Histone H1 type 4-A (HIST1H4A) (Fig. 4B). Expression of SOD1 increased with pioglitazone treatment and it was found to interact with 2 other upregulated proteins, PARK7 (parkinsonism associated deglycase) and CCS (copper chaperone for superoxide dismutase). Interestingly, downregulation of BMI1 and its interacting histone proteins in the network model suggests potential epigenetic regulation by pioglitazone, which is consistent with functional enrichment data indicating “chromatin silencing” (Fig. 3A).
Biological Influences of Pioglitazone on Normal Bladder Epithelial Cells
To further validate the DEPs between the control versus pioglitazone groups, proteins were selected for based on functional enrichment analyses. Western blot analysis showed that expression of MYH3 was upregulated, while expression of ACTG2 was downregulated in cells treated with 25μM of pioglitazone (Fig. 5A). Consistent with morphological changes in pioglitazone-treated cells, we noticed that protein expression of mesenchymal cell markers (N-cadherin, Snail, Slug, and β-catenin) were downregulated in response to pioglitazone treatment (Fig. 5B). In addition, tight junction proteins (zonula occludens [ZO] 1 and 2 and CD2-associated protein) that are involved in cell-cell contact regions were greatly reduced in pioglitazone-treated cells (Fig. 5C).
To determine whether pioglitazone regulates mitochondrial quantity or metabolic respiration-associated proteins, mitochondrial OXPHOS proteins were measured via western blot analysis. The expression levels of OXPHOS subunits (C I subunit [NDUFB8], C II subunit [succinate dehydrogenase, SDHB], C III core protein 2 [ubiquinol-cytochrome C reductase core protein 2, UQCRC2], C IV subunit [mitochondrially encoded cytochrome C oxidase I, MTCO1], and C V alpha subunit [ATP synthase subunit alpha, ATP5A]) were not change by pioglitazone treatment (Fig. 5D). To profile the phosphorylation events in normal bladder cells, human TRT-HU1 cells were treated with or without pioglitazone. A series of western blot analyses showed that several phosphorylation events of specific signaling pathways were regulated by pioglitazone. The phosphorylation levels of NF-κB, Erk/MAPK, ERBB2, and PAK1 decreased. On the contrary, only the phosphorylation levels of GSK-3β increased. Additionally, previous studies have shown that the activation of PPARγ through agonists reduces diabetic myocardial fibrosis by transforming growth factor beta/extracellular signal-regulated kinase pathway and epithelial-mesenchymal transition [16], which were both found to be upregulated in pioglitazone-treated cells compared to controls (Fig. 5E).
To further elucidate on the role of pioglitazone on oxidative stress, ROS levels were measured and analyzed after 72-hour pioglitazone treatment. Levels of ROS were unchanged between the control and pioglitazone groups (Fig. 5F). In addition to our proteomic analysis, we assessed whether pioglitazone affects cell proliferation. The dose-dependent experiments in varying concentrations of pioglitazone (25, 50, or 100μM) for 24 and 48 hours, we noticed that the cell proliferation rates of cells treated for 48 hours were significantly suppressed, compared to controls (Fig. 5G, H).
DISCUSSION
There are many discrepancies surrounding the link between TZD usage and side effects on bladder function. Diabetic bladders can have several dysfunctions, ranging from bladder overactivity to impaired contractility, which represent a wide spectrum of clinical symptoms. The widespread pathophysiology of bladder dysfunction in DM patients is associated with multiple factors like smooth muscle dysfunction, and urothelial impairment [17]. DM has been shown to be a significant predictor of death from cancer, including BC [18]. Evidence from a metaanalysis study has linked the use of pioglitazone to increased BC risk as well [13]. The tumorigenic effect of pioglitazone is thought to be due to signaling modifications in the PPAR pathway [19]. However, it is still unclear whether the long-term use of pioglitazone increases the chances of developing bladder dysfunctions or disease.
PPARγ is part of the PPAR subfamily that consists of 2 other subunits, alpha and beta/delta. As the most extensively studied subunit, the functions of PPARγ have been linked to several crucial biological processes, such as glucose metabolism, lipid biosynthesis, and anti-inflammatory response [20]. The association of PPARγ to diabetes is based on its role in lipid and glucose metabolism [21]. It has been shown that patients with dominant-negative mutations of PPARγ have severe hypoglycemia, insulin resistance, and DM [22]. This means that there is a significant genetic link between both PPARγ and DM. Treatment with TZDs has proven to substantially decrease insulin resistance in diabetes and possibly induces transcription of adipose cell differentiation, which generates smaller insulin-sensitive adipocytes [23]. However, despite their beneficial effects in patients with diabetes, TZDs are known to have adverse effects. This includes weight gain, fluid retention, bone mineral density reduction, and excess myocardial infarction [24]. The fluid retention induced by TZDs has also been proven to significantly increase urine volume and excretion of sodium [25]. Because of this, there is some controversy surrounding the safety of long-term TZD usage to treat diabetes.
In addition to the documented roles of PPARγ in lipid metabolism and DM, there has been growing research on its functions in both BC and interstitial cystitis (IC). Approximately 20%–25% of human BC is driven by PPAR signaling, PPARγ gene amplification, or RXRA hot-spot mutations [26]. Furthermore, it was found that the PPARγ transcription pathway associated with an immune evasion in muscle-invasive BC [27]. Along with this immune escape, it has been proposed that PPARγ cooperates with the FOXA1 (forkhead box protein A1) and GATA3 (GATA binding protein 3) networks to drive transdifferentiation of basal BC cells into luminal types [28]. PPARγ has also been identified to be associated with IC. Mahal et al. [29] found that treating their IC rat model with PPARγ agonists improved bladder function and urinary health. Based on the complex relationship of PPARγ with diabetes and urological malignancies, it would be compelling to better understand how patients who routinely take TZDs are affected.
This study revealed that BMI1 is a master node in pioglitazone-regulated networks. Our proteomic profiling of BMI1 exhibited significant downregulation in normal bladder epithelial cells treated with pioglitazone, suggesting a potential inverse relationship between PPARγ and BMI1. One possible hypothesis for our future study is that PPARγ signaling induced by pioglitazone may cause loss of BMI1 expression in bladder cells. Expression of telomere maintenance proteins including HIST1H4A, HIST2H2AA3, and HIST2H2AC, which interact with BMI1, are downregulated as well. The presence of an upregulated telomere maintenance mechanism is a hallmark of cancer, and it is interesting to note that HIST2H2AA3 and HIST2H2AC were identified as genes upregulated, differentially expressed in BC [30]. This may signify a stage shift, by pioglitazone treatment, from developing cancer. PPARγ agonism has also been shown to increase the antitumor effects of histone deacetylase inhibitors in multiple myeloma cells [25].
Most notably, in our study, we found a series of contractile and cytoskeleton proteins, such as myosin light polypeptides, myosin heavy polypeptides, keratins, and ACTG2, as being significantly up- or downregulated in TRT-HU1 bladder epithelial cells treated with pioglitazone. Myosin is a particularly important structural component of muscles that interacts with actin filaments and plays a vital role in actin filament contraction, bundling, motor action, and binding. Furthermore, our network modeling revealed that these actin-binding proteins comprise the central hub. These results suggest the potential pathophysiological mechanisms of pioglitazone’s effects.
Our data demonstrated that the activation of PPARγ signaling pathways suppressed the proliferation of bladder epithelial cells. Interestingly, these findings are consistent with clinical observations showing that the IC bladder has thinner epithelial layers [31] and that IC bladder cells exhibit reduced proliferation due to cell cycle arrest [32]. Our epigenetic remodeling study supports this mechanistic link between IC and defective cell proliferation [15], Furthermore, previous reports suggest that the antiproliferative role of PPARγ is through the upregulation of PTEN expression in various cancers, which leads to decreased PI-3K activity [33]. It is currently known that PPARγ agonists suppress cell cycling by downregulating cyclin D1 in several cancer cells; thereby, inhibiting cyclin D1/CDK-mediated retinoblastoma (pRb) phosphorylation and maintaining its active form to prevent the G1 to S phase transition [34]. Activation of PPARγ via agonists in normal human urothelial cells has been shown to link with late/terminal urothelial differentiation by protein induction of uroplakins, cytokeratins, and tight junction constituents [35].
We very much understand certain limitations of the study design arising from performing experiments in normal bladder cells, which cannot truly reflect the diabetic bladder condition [36,37]. Although additional experiments using bladder cells from diabetic condition are actually going on in our laboratory, it is out of scope of this study aiming to understand the pioglitazone effects on normal bladder cells. Another limitation of this study is that we used only one normal bladder cell line. This present study also suffers from limited impact on clinical application mainly due to in vitro experiments.
In summary, although pioglitazone is widely used among T2DM patients, its effects on the bladder are still in controversy. While there have been studies examining the effects of pioglitazone on healthy bladder cells, there is no conclusive evidence. Through unbiased quantitative proteomic profiling, our report comprehensively characterized the proteome of normal bladder epithelial cells treated with pioglitazone. We observed suppressed cell proliferation, which may be explained by the decreased expression of tight junction proteins. Taken together, our study provides no experimental evidence to support a tumorigenic effect of pioglitazone on the bladder. This proteomic approach will be useful for examining any potential side effects of the one of the most commonly used drugs.