ITRODUCTION
The normal function of the lower urinary tract (LUT) requires coordination of the urinary bladder, urethra, pelvic floor, efferent and afferent neurons, and specific spinal cord and brain areas [1]. The bladder and external urethral sphincter are innervated directly or indirectly by many central nervous system (CNS) regions, such as the pontine micturition center (PMC), locus coeruleus, hypothalamus, and preoptic area [2]. The PMC, as the supraspinal switching center, is known to regulate the storage and elimination of urine [3]. The periaqueductal gray matter (PAG) of the hypothalamus has also been suggested to be a critical component of the micturition reflex, and interneurons in the lumbosacral cord strongly project to the lateral and dorsal parts of the PAG [4].
The central micturition regions, including the PAG and PMC, are activated by stimulation of the bladder. Many studies have reported that bladder stimulation caused the expression of c-Fos in the PAG and PMC related to micturition [3,5]. The transcription factor c-Fos, an immediate early gene, has been used as a marker of neuronal activity [6]. Nerve growth factor (NGF), the prototypical neurotrophic factor, has also been reported to modulate neuronal function along micturition pathways, and to serve as a urinary marker of bladder overactivity [7].
Nitric oxide (NO) is considered to be an important neurotransmitter and cell signaling molecule with a broad range of functions in the LUT. NO has been reported to play an important role in the micturition process and in disorders of the LUT [8]. NO is synthesized from L-arginine by nitric oxide synthase (NOS), which is classified into 3 groups: neuronal NOS (nNOS), inducible NOS, and endothelial NOS. Zhang et al. [9] reported that NO participates in the neural pathways controlling the LUT, and that NOS alterations may be involved in LUT dysfunction.
As described above, the functions of the LUT, such as voiding and storing urine, are dependent on neural circuits located in the brain, spinal cord, and peripheral ganglia. Therefore, the functions of the LUT are susceptible to various neurologic disorders including spinal cord injury (SCI) [10]. SCI may result from traumatic insult, such as spinal transection or contusion, or ischemic insult occurring at different levels of the spinal cord [11], and these injuries generate a range of events compromising the integrity of the neuronal pathways running through the spinal cord [12]. Virtually, SCI at the cervical or thoracic levels disrupts voluntary control of voiding and the normal reflex pathways that coordinate bladder and sphincter functions [13]. Thus, SCI animal models have been used to study the mechanism of detrusor overactivity and detrusor-sphincter dyssynergia [14].
The most commonly used first-line drugs for male patients with lower urinary symptoms are α1-adrenoceptor blockers, such as silodosin and prazosin, which relieve voiding symptoms and storage symptoms in elderly men with BPH by exerting an inhibitory effect on afferent input from the LUT [15].
Tamsulosin, an α1-adrenoceptor blocker, is also considered the most effective regimen for patients with LUT symptoms suggestive of benign prostatic hyperplasia, and has been reported to play a role in the management of overactive bladder (OAB) in women [16]. In addition, tamsulosin has been reported to improve storage function by inhibiting C-fiber mediated urethral afferent neurons [17]. Abrams et al. [18] studied the efficacy and safety of tamsulosin in patients with suprasacral spinal cord lesions, and suggested that long-term tamsulosin treatment improves bladder storage and emptying, alleviating the symptoms of autonomic dysreflexia. However, they did not propose an exact mechanism of the effects of tamsulosin on the afferent micturition regions. Therefore, the effects of tamsulosin on the expression of c-Fos, NGF and nicotinamide adenine dinucleotide phosphate-diaphorase (NADPH-d) in the afferent micturition areas, such as the PMC, the ventrolateral PAG (vlPAG), and the spinal cord (L5), of rats with an SCI were investigated in this study.
MATERIALS AND METHODS
Animals
Female Sprague-Dawley rats weighing 260±10 g (13 weeks of age) were used in this study. All experimental procedures were carried out in accordance with the Guidelines for the Care and Use of Animals approved by the Council of the National Institutes of Health Guide for the Care and Use of Laboratory Animals. The study was approved by the Institutional Care and Use Committee of Kyung Hee University (KHUASP[SE]-17-093). To minimize inadvertent c-Fos protein expression induced by environmental stimuli, the animals were gently and carefully handled during the experimental process. The animals were housed under controlled temperature (22°C±2°C) and lighting (8 AM to 8 PM) conditions and were supplied with food and water ad libitum.
The animals were randomly divided into the following 4 groups (n=8 in each group): the sham group, the SCI-induced group, the SCI-induced and 0.01 mg/kg tamsulosin-treated group, and the SCI-induced and 1 mg/kg tamsulosin-treated group. Starting one day after the induction of SCI, the tamsulosin-treated groups received tamsulosin (Harunal, Astellas Pharma Inc., Tokyo, Japan) orally once a day for 3 weeks.
Surgical Procedure for Inducing SCI
The SCI model was induced as previously described [19]. The rats were anesthetized by inhalation of isoflurane (2% isoflurane in 30% O2 and 70% N2, JW pharmaceutical, Seoul, Korea) during surgery. The skin in the T10–T12 areas was incised through a 2.5-cm median incision, and the thoracic vertebral column and spinous process in the T11 were exposed by dissection. After dissection, the spinous process in T11 was ground using a drill until the vertebral arch area was reached, and a hole 2 mm deep from the surface of the vertebral arch was made using a surgical drill (diameter, 1 mm), the spinal cord was damaged by a 22-G needle, and the hole was sealed (Fig. 1). The skin was closed layer by layer with continuous 3-0 silk sutures. The body temperature was maintained at 36°C ±0.5°C during the induction of SCI using a Homeothermic Blanket Control Unit (Harvard Apparatus, Holliston, MA, USA) that enveloped the body and the head. After recovery, the animals were monitored for an additional 2 hours to prevent hypothermia. The animals in the sham-operation group were treated identically, except that the spinal cords was not damaged after the skin incisions.
Cystometry
Bladder function was evaluated using cystometry, as previously described [20]. The rats were tested via cystometry after 3 weeks of tamsulosin treatment. The rats were anesthetized with Zoletil 50 (10 mg/kg, intraperitoneally; Vibac Laboratories, Carros, France). A sterile polyethylene catheter (PE50) was inserted into the urethra through the bladder dome. The catheter was connected to a pressure transducer (Harvard Apparatus) and syringe pump (Harvard Apparatus) via a 3-way stopcock to record the intravesical pressure and to infuse saline into the bladder. After the bladder was emptied, cystometry was performed with saline infused at 0.5 mL. The basal contraction pressure and contraction time in the bladder were monitored using Labscribe software (iWorx/CB Science Inc., Dover, DE, USA).
Tissue Preparation
The animals were sacrificed immediately after determining their voiding function by cystometry as previously described [19,20]. For the histological examination, the rats were transcardially perfused with 50mM phosphate-buffered saline (PBS), followed by 4% paraformaldehyde in 100mM sodium phosphate buffer at pH 7.4. The brain was removed, postfixed in the same fixative overnight, and transferred into a 30% sucrose solution for cryoprotection. Serial 40-μm-thick coronal sections were made with a freezing microtome (Leica, Wetzlar, Germany). The PMC was selected from the region spanning from Bregma –9.68 to –9.80 mm. The vlPAG was selected from the region spanning from Bregma –7.64 to –8.00 mm, and the spinal cord was selected from the region spanning L5. Four sections on average in each region were collected from each rat.
Immunohistochemistry for c-Fos and NGF
To detect c-Fos expressions in the vlPAG, PMC, and spinal cord (L5) regions, c-Fos and NGF immunohistochemistry was performed as previously described [20]. Briefly, tissue sections were incubated overnight with rabbit anti-c-Fos and mouse anti-NGF-antibodies (Santa Cruz Biotechnology Inc., Santa Cruz, CA, USA) at a dilution of 1:1,000. The sections were then incubated for 1 hour with anti-rabbit and anti-mouse secondary antibodies (1:200; Vector Laboratories, Burlingame, CA, USA). The sections were subsequently incubated with an avidin–biotin–peroxidase complex (1:100; Vector Laboratories) for 1 hour at room temperature. Immunoreactivity was visualized by incubating the sections in a solution consisting of 0.02% 3,3´-diaminobenzidine tetrahydrochloride (Sigma-Aldrich, St. Louis, MO, USA) and 0.03% H2O2 in 50mM Tris–HCl (pH, 7.6). The sections were then washed 3 times with PBS and mounted onto gelatin-coated slides. The slides were air-dried overnight at room temperature, and the coverslips were mounted using Permount (Fisher Scientific, Waltham, MA, USA).
Histochemistry for NADPH-d
To detect NOS activity in the PMC, vlPAG, and spinal cord (L5) regions, NADPH-d histochemistry was performed as previously described [20]. Briefly, free-floating sections were incubated at 37ºC for 60 minutes in 100mM PBS (pH, 7.4) containing 0.3% Triton X-100, 0.1 mg/mL of nitroblue tetrazolium, and 0.1 mg/mL of β-NADPH. The sections were then washed 3 times with PBS and mounted onto gelatin-coated slides. The slides were air-dried overnight at room temperature, and the coverslips were mounted using Permount (Fisher Scientific).
Data Analysis
To assess c-Fos, NGF and NADPH-d expressions in the neuronal afferent pathways of micturition (vlPAG, PMC, and spinal cord L5), cell counting was performed through a light microscope (BX-51; Olympus Corp., Tokyo, Japan). The area of the neuronal afferent pathways of micturition in each slice was measured using the Image-Pro Plus computer-assisted image analysis system (Media Cyberbetics Inc., Silver Spring, MD, USA) attached to the light microscope (BX-51; Olympus Corp.). The numbers of cells positive for c-Fos, NGF, and NADPH-d in the vlPAG and PMC were counted hemilaterally, and the numbers of cells positive for c-Fos, NGF, and NADPH-d in the spinal cord L5 were counted on both sides of the dorsal horn. The results were expressed as the number in the selected region per section.
Statistical analysis was performed using IBM SPSS Statistics ver. 20.0 (IBM Co., Armonk, NY, USA). The data are expressed as the mean±standard error of the mean. For comparisons between groups, 1-way analysis of variance and the Duncan post hoc test were performed, and P-values <0.05 were considered to indicate statistically significant differences among the groups.
RESULTS
Effects of Tamsulosin on Basal Contraction Pressure and Time According to Cystometry
The basal contraction pressure and time according to cystometry are presented in Fig. 2. The basal contraction pressure and time in the SCI-induced group were significantly increased compared to the sham group (P<0.05). In contrast, tamsulosin treatments significantly decreased the basal contraction pressure and time, dose-dependently. In particular, treatment with 1 mg/kg of tamsulosin reduced the basal contraction pressure to a level similar to that of the sham group.
Effects of Tamsulosin on c-Fos Positive Cells in the vlPAG, PMC, and Dorsal Horn of the Spinal Cord (L5)
Photomicrographs of c-Fos-positive cells in the vlPAG, PMC, and dorsal horn of the spinal cord (L5) are presented in Fig. 3. These data showed that the expression of c-Fos in the vlPAG, PMC, and spinal dorsal horn (L5) of the SCI-induced group was conspicuously increased compared to the sham group (P<0.05). However, tamsulosin treatment significantly suppressed the expression of c-Fos in the vlPAG, PMC, and spinal dorsal horn (L5) in comparison to the SCI-induced group (P<0.05), and the inhibitory effect of tamsulosin on the expression of c-Fos in the spinal cord was dose-dependent.
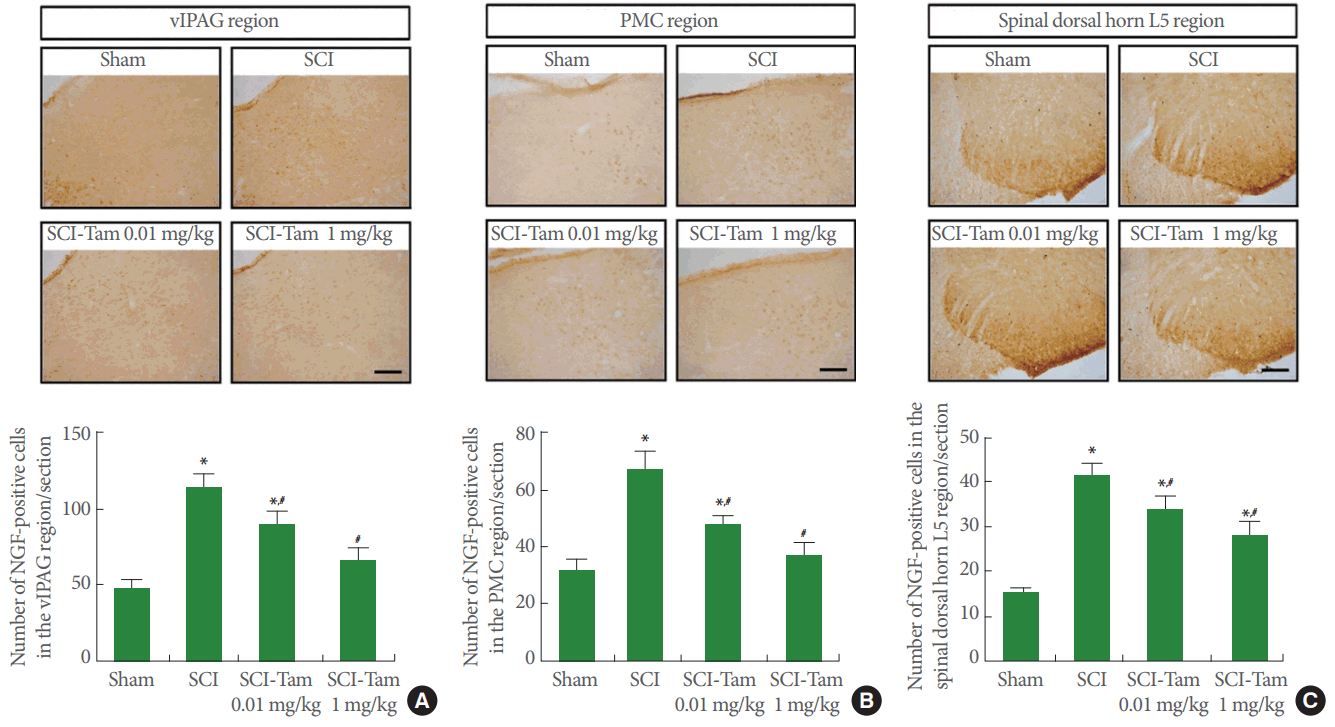
Effects of Tamsulosin on NGF-Positive Cells in the vlPAG, PMC, and Dorsal Horn of the Spinal Cord (L5)
Photomicrographs of NGF-positive cells in the vlPAG, PMC, and spinal dorsal horn (L5) are presented in Fig. 4. Similar to the expression of c-Fos, the expression of NGF in the vlPAG, PMC, and spinal dorsal horn (L5) of the SCI-induced group was remarkably increased, compared to the sham group (P<0.05). Moerover, tamsulosin treatment significantly suppressed the expression of NGF in the vlPAG, PMC, and spinal dorsal horn (L5) compared to the SCI-induced group (P<0.05). This suppression of NGF expression in the vlPAG, PMC, and dorsal horn of the spinal cord by tamsulosin was proportional to the doses of these agents.
Effects of Tamsulosin on NADPH-d Positive Cells in the vlPAG, PMC, and Dorsal Horn of the Spinal Cord (L5)
Photomicrographs of NADPH-d-positive cells in the vlPAG, PMC, and dorsal horn of the spinal cord (L5) are presented in Fig. 5. While the expression of NADPH-d in the vlPAG, PMC, and spinal dorsal horn (L5) of the SCI-induced group remarkably increased, tamsulosin treatment significantly suppressed the expression of NADPH-d in the vlPAG, PMC, and spinal dorsal horn compared to the SCI-induced group (P<0.05). Tamsulosin treatment reduced the number of NADPH-d-positive cells in the vlPAG, PMC, and spinal dorsal horn to the level of the sham group.
DISCUSSION
Among the many experimental SCI models that have been developed, weight drop methods and the clip-damage model after laminectomy resulted in low survival rates due to various side effects after surgery. Therefore, in this study, SCI was induced by the method of Cho et al. [19]. The advantage of this model is that it has a high survival rate of animals because of the rapid operation and because the affected part is minimized.
SCI impairs the neuronal pathways running through the spinal cord, and animal models of SCI induced by chemical lesions, induction of ischemia, and complete or incomplete transection are used to study pathologies related to SCI, including bladder dysfunction [11,12]. SCI has been reported to induce substantial anatomical reorganization of spinal circuits, to affect the imbalance between excitatory and inhibitory inputs, and to impair certain cellular functions [21]. Consequently, SCI significantly impairs micturition and results in areflexic detrusor underactivity and neurogenic detrusor overactivity, depending the level of the lesion [12]. de Groat and Yoshimura [13] reported that SCI at the cervical or thoracic levels disrupted voluntary control of voiding and the normal reflex pathways that coordinate bladder and sphincter functions. Miyazato et al. [14] also reported that SCI induced by transection of T9–10 significantly increased non-voiding bladder contractions and decreased bladder capacity. Similarly, the present study found that SCI induced by incomplete transection of T11 significantly increased the basal contraction pressure and time. Such increases in the basal contraction pressure and time have been found in cyclophosphamide-induced OAB and the spontaneous hypertensive rat animal model of detrusor overactivity and OAB [20,22].
Previous studies suggested that these afferent signals and their central processing play an important role in normal LUT function and in the pathogenesis of functional LUT diseases, such as OAB and neurogenic bladder [11-13,17]. Following SCI, the bladder is initially a reflexic but then becomes hyperreflexic, due to the emergence of a spinal micturition reflex pathway [13]. Sensory afferent fibers run in the 3 sets of peripheral nerves—pelvic, hypogastric, and pudendal—and transmit information to the CNS about bladder filling. Most of the bladder sensory fibers run in the pelvic nerve, and their cell bodies are located in the sacral dorsal root ganglia in humans and in L5–S1 in rats [12]. These sacral afferent fibers send their projections to the PAG, via the supraspinal pathway [4]. The bladder information is first processed within the PAG, and then it is relayed to the PMC. The PMC, as the supraspinal switching center, sends excitatory motor input to the sacral spinal cord, resulting in detrusor contraction and relaxation of the urethra [3,23]. More specifically, the bladder, via the lumbosacral spinal cord, sends information about bladder filling to the PAG, and then PMC-projecting cells in the PAG become activated and initiate micturition.
Afferent fibers from the bladder have been reported to express tropomyosin-related kinase A, a high-affinity receptor for NGF [24]. NGF has been reported to modulate neuronal function associated with micturition and to be involved in the development of neurogenic bladder overactivity at the spinal level [25]. Yoshimura et al. [26] reported that increased NGF levels in bladder afferent pathways and NGF-induced reduction in A-type K+ current density could contribute to the hypertrophy and hyperexcitability of bladder afferent neurons, as well as to the emergence of bladder overactivity. Our results also showed that the expression of NGF in the spinal dorsal horn of the SCI-induced group was conspicuously increased in comparison with the sham group, and, similarly to NGF expression in the lumbosacral spinal cord, NGF expression was upregulated in the vlPAG and PMC. The levels of NGF are strongly enhanced in SCI animals, and alterations in the synthesis and release of neurotrophic factors in the peripheral and central nervous system could contribute to the emergence of the spinal micturition reflex [27].
de Groat and Yoshimura [10] suggested that the expression of c-Fos in the spinal cord identifies that the spinal neurons are involved in processing afferent input from the LUT. Such an increase of neuronal activities in the lumbosacral spinal cord may stimulate the supraspinal pathways regulating micturition. In fact, many studies have reported that stimulation of the bladder caused the expression of c-Fos in the PAG and PMC [3,5]. Microinjections of excitatory neurotransmitters, such as glutamate, acetylcholine, or γ-aminobutyric acid antagonist, into the PMC induce bladder contraction, decreases the threshold bladder volume causing the micturition reflex, or increase the amplitude of bladder contraction [28]. Based on these studies, it can be suggested that stimulation or enhanced neuronal activity in the PMC and PAG might be involved in the symptoms of OAB. In the present results, the expression of c-Fos in the spinal dorsal horn, vlPAG, and PMC in the SCI-induced group was significantly upregulated compared to the sham group, and the basal contraction pressure and time were greater in the SCI-induced group than in the sham group.
SCI can induce the upregulated expression of NOS, and of particular note, the upregulation of nNOS may facilitate the emergence of the spinal micturition reflex following SCI [9]. Kakizaki and de Groat [29] suggested that NO at the spinal level participates in the facilitation of the micturition reflex by nociceptive bladder afferent neurons activated by noxious chemical irritation of the bladder. Lagos and Ballejo [30] also reported that the increased production of NO in spinal L6–S2 segments seems to be necessary for the initiation of micturition hyperreflexia based on the identification of increased Ca2+-dependent NOS activity in the micturition hyperreflexia observed in the initial hours of cyclophosphamide-induced cystitis. Neurons containing NO have been identified histochemically by the presence of NADPH-d [31], and such NADPH-d activity has also been identified in neurons in the spinal cord [29]. The present study also showed that the expression of NADPH-d in the spinal dorsal horn was conspicuously upregulated following SCI, suggesting an increased production of NO. In addition, upregulation of NADPH-d occurred in the vlPAG and PMC, the supraspinal afferent pathways.
As shown in this study, SCI upregulated neural activities and the production of NO in the sacral afferent pathway (L5) and the supraspinal pathways, the vlPAG and PMC, resulting in OAB. However, tamsulosin treatment significantly suppressed the expression of c-Fos and NGF in the dorsal horn of the spinal cord (L5), the vlPAG, and the PMC. Moreover, tamsulosin treatment remarkably reduced the expression of NADPH-d in the afferent pathways of micturition. Consequently, tamsulosin significantly attenuated the symptoms of OAB.
Activation of the α1-adrenoceptor in the bladder causes OAB symptoms [32]. Thus, tamsulosin, an α1-adrenoceptor antagonist, has been reported to have beneficial effects on the micturition function [20,22]. Yokoyama et al. [17] reported that tamsulosin had an inhibitory effect on the C-fiber urethral afferent nerves, resulting in improved bladder storage. Most sensory afferent nerves innervating the LUT, especially the bladder, are small myelinated (Aδ fibers) or unmyelinated (C fibers) axons. In particular, C fibers can be identified by their neuropeptide content and expression of transient receptor potential vanilloid 1 [33]. In chronic SCI rats, C-fiber afferent neurons mediate bladder hyperreflexia and automatic micturition, leading to detrusor-sphincter dyssynergia, which refers to simultaneous contractions of the external urethral sphincter and bladder during the micturition reflex [13,34].
Seki et al. [34] reported that increased levels of NGF in the spinal cord are involved in the emergence of detrusor-sphincter dyssynergia which is partially mediated by C-fiber bladder afferent neurons after SCI. NGF has been reported to induce hyperexcitability in C-fiber bladder afferent pathways and bladder hyperactivity in various pathological conditions, including SCI [27,34]. Yoshimura et al. [26] also reported that increased NGF levels in bladder afferent pathways and NGF-induced reduction in A-type K+ current density could contribute to the hyperexcitability of bladder afferent neurons and bladder overactivity. However, neutralization of NGF in the spinal cord suppressed C-fiber mediated bladder hyperreflexia in SCI-induced rats [34]. Thus, suppression of NGF levels in afferent pathways could be useful for treating detrusor-sphincter dyssynergia or voiding dysfunction following SCI [10,34]. In the present study, tamsulosin treatment also significantly suppressed the expression of NGF in the dorsal horn of the spinal cord (L5) in SCI-induced rats, and this inhibitory effect of tamsulosin on NGF expression also occurred in the supraspinal pathways, the vl-PAG, and the PMC. In addition, tamsulosin suppressed the expressions of c-Fos in the spinal cord (L5), the vlPAG and the PMC. Suppressing the expression of c-Fos in afferent pathways regulating micturition contributes to the attenuation of OAB symptoms, and this inhibitory effect of tamsulosin has been found in other studies [20,22].
Finally, tamsulosin suppressed the expression of NADPH-d in the dorsal horn of the spinal cord (L5), the vlPAG, and the PMC to a remarkable extent, suggesting that NO production was suppressed in these regions. C-fiber afferent neurons are sensitive to NO, and the increased production of NO may be necessary for the initiation of micturition hyperreflexia [30]. Moreover, the production of NO seems to be associated with increased spinal Fos expression [35]. Lagos and Ballejo [30] found that the nonselective NOS inhibitor NG-nitro-L-arginine-methyl ester diminished the hyperreflexic state when administered systemically, and Zhang et al. [9] also reported that an nNOS inhibitor suppressed SCI-induced urinary incontinence by increasing bladder capacity.
The results of the present study demonstrate that SCI increased nonvoiding bladder contractions and reduced bladder capacity by increasing the expression of NGF and c-Fos and NO production in the afferent pathways of micturition, which suggests that the bladder afferent neurons became hyperexcitable and that OAB was induced. However, tamsulosin treatment showed beneficial effects on alleviating the symptoms associated with bladder hyperactivity by suppressing hyperexcitability in the afferent pathways of micturition (lumbosacral spinal cord, the vlPAG, and the PMC). Therefore, tamsulosin, an α1-adrenoceptor antagonist, may be able to attenuate bladder dysfunction following SCI. However, this study did not investigate the other regions involved in micturition, such as the anterior cingulate gyrus, medial preoptic nucleus, and frontal cortex. Thus, further studies are needed to elucidate the exact mechanism and role of tamsulosin on the afferent pathways of micturition.