Rocuronium Bromide Inhibits Inflammation and Pain by Suppressing Nitric Oxide Production and Enhancing Prostaglandin E2 Synthesis in Endothelial Cells
Article information

Abstract
Purpose
Rocuronium bromide is a nondepolarizing neuromuscular blocking drug and has been used as an adjunct for relaxation or paralysis of the skeletal muscles, facilitation of endotracheal intubation, and improving surgical conditions during general anesthesia. However, intravenous injection of rocuronium bromide induces injection pain or withdrawal movement. The exact mechanism of rocuronium bromide-induced injection pain or withdrawal movement is not yet understood. We investigated whether rocuronium bromide treatment is involved in the induction of inflammation and pain in vascular endothelial cells.
Methods
For this study, calf pulmonary artery endothelial (CPAE) cells were used, and 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide assay, Western blot, nitric oxide detection, and prostaglandin E2 immunoassay were conducted.
Results
Rocuronium bromide treatment inhibited endothelial nitric oxide synthase and suppressed nitric oxide production in CPAE cells. Rocuronium bromide activated cyclooxygenase-2, inducible nitric oxide synthase and increased prostaglandin E2 synthesis in CPAE cells.
Conclusions
Rocuronium bromide induced inflammation and pain in CPAE cells. Suppressing nitric oxide production and enhancing prostaglandin E2 synthesis might be associated with rocuronium bromide-induced injection pain or withdrawal movement.
INTRODUCTION
Rocuronium bromide is a nondepolarizing neuromuscular blocking drug, and it has been used as an adjunct for relaxation or paralysis of skeletal muscles, facilitation of endotracheal intubation, and improving surgical conditions during general anesthesia [1,2]. The introduction of rocuronium bromide to the anesthetic field replaced succinylcholine for rapid sequence intubation because of its rapid onset time and intermediate duration. Rocuronium bromide avoids the complications of succinylcholine; however, injection pain or withdrawal movement has been reported. Injection pain or withdrawal movement is characterized by sudden flexion of the wrist and arm after rocuronium infusion, lasting 10–20 seconds [1]. The incidence of rocuronium-induced injection pain or withdrawal movement is reportedly 50%–80%. The incidence and severity of injection pain or withdrawal movement are higher in children than in adults [3]. Activation of nociceptors by the osmolality or pH of solution and release of endogenous inflammatory mediators, such as histamine, kinin, and other substances, have been suggested as the underlying mechanisms of rocuronium bromide-induced injection pain or withdrawal movement [1,4,5]. However, the exact mechanism of rocuronium bromide-induced injection pain or withdrawal movement is not well understood.
Prostaglandin E2 (PGE2) is a key inflammatory mediator that is synthesized from arachidonic acid via a cyclooxygenase (COX)-dependent pathway. There are 2 isoforms of COX: COX-1 and COX-2. While COX-1 is a constitutively expressed form under normal physiological functions, COX-2 is expressed only in response to inflammatory signals, such as cytokines and bacterial endotoxin lipopolysaccharide (LPS), in inflammatory cell types such as fibroblasts, monocytes, and vascular endothelial cells. COX-2 produces PGE2, which induces inflammation [6,7].
Nitric oxide (NO) is a ubiquitous free radical that plays an important role either as a messenger or destructive molecule in inflammation, and NO modulates the inflammatory process [8]. NO is endogenously generated from L-arginine by NO synthase (NOS), and NO is implicated in many physiological processes [9-11]. Several isoforms of NOS exist, and these isoforms fall into three major classes: inducible NOS (iNOS), endothelial NOS (eNOS), and neuronal NOS. Of these, iNOS and eNOS are important enzymes in the regulation of inflammation [9]. iNOS is highly expressed in macrophages, neutrophils, and endothelial and smooth muscle cells in response to different stimuli [12]. Endothelium-derived NO is synthesized by eNOS from the precursor L-arginine, and it is not only a major mediator of endothelium-dependent vasodilation but also critically involved in the protective property of the healthy endothelium [13].
In the present study, we investigated whether rocuronium bromide is involved the induction of inflammation and pain in vascular endothelial cells. For this, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay, Western blot for COX-1, COX-2, 5-LOX, iNOS, eNOS, and NO detection, and PGE2 immunoassay were conducted in calf pulmonary artery endothelial (CPAE) cells after rocuronium bromide treatment.
MATERIALS AND METHODS
Reagents
Rocuronium bromide was obtained from Korea Organon (Seoul, Korea). LPS was obtained from Sigma Chemical Co. (St. Louis, MO, USA). MTT assay kit was purchased from Boehringer Mannheim (Mannheim, Germany).
Cell Culture
CPAE cells were purchased from the Korean Cell Line Bank (KCLB, Seoul, Korea). CPAE cells were cultured in Roswell Park Memorial Institute (RPMI) 1640 medium (Gibco BRL, Grand Island, NY, USA) supplemented with 10% heat-inactivated fetal bovine serum (FBS; Gibco BRL) at 37°C in 5% CO2–95% O2 in a humidified cell incubator. The cells were plated onto culture dishes at a density of 2×104 cells/cm2 , 24 hours prior to drug treatments.
MTT Cytotoxicity Assay
CPAE cells were grown in 100 μL of culture medium per well in 96-well plates. To determine the cytotoxicity of rocuronium bromide, MTT assay was conducted according to the previously described method [14]. The cells were treated with rocuronium bromide at concentrations of 1, 5, 10, 50, 100, 500, and 1,000 μg/mL for 24 hours. The cells in the control group were left untreated. After adding 10 µL of MTT labeling reagent containing 5 mg/mL MTT in phosphate-buffered saline to each well, the plates were incubated for 2 hours. Next, 100 µL of solubilization solution containing 10% sodium dodecyl sulfate (SDS) in 0.01M hydrochloric acid was added to each well, and the cells were incubated for another 12 hours. The absorbance was then measured with a microtiter plate reader (Bio-Rad, Hercules, CA, USA) at a test wavelength of 595 nm with a reference wavelength of 690 nm. The optical density (O.D.) was calculated as the difference between the absorbance at the reference wavelength and that observed at the test wavelength. Percent viability was calculated as follows: (O.D. of drug-treated sample/control O.D.) ×100.
Western Blot Analysis
The expression levels of COX-1, COX-2, iNOS, and eNOS were determined by Western blot analysis according to the previously described method [15]. CPAE cells were lysed in an ice-cold whole cell lysate buffer containing 50mM HEPES (N-2-hydroxyethylpiperazine-N-2-ethanesulfonic acid) (pH, 7.5), 150mM NaCl, 10% glycerol, 1% Triton X-100, 1.5mM magnesium chloride hexahydrate, 1mM EGTA (ethyleneglycol-bis-(β-aminoethyl ether)-N,N´-tetraacetic acid), 1mM phenylmethylsulfonyl fluoride, 2-μg/mL leupeptin, 1-μg/mL pepstatin, 1mM sodium orthovanadate, and 100mM sodium fluoride, and the mixture was incubated for 30 min at 4°C. Cell debris was removed by microcentrifugation, followed by quick freezing of the supernatant.
The protein concentration was measured using a Bio-Rad colorimetric protein assay kit (Bio-Rad). Thirty micrograms of protein was separated on SDS-polyacrylamide gels and transferred onto a nitrocellulose membrane (Schleicher & Schuell GmbH, Dassel, Germany). Goat COX-1 antibody (1:2,000; Santa Cruz Biotechnology, Santa Cruz, CA, USA), goat COX-2 antibody (1:2,000; Santa Cruz Biotechnology), rabbit iNOS antibody (1:500; Santa Cruz Biotechnology), and goat eNOS antibody (1:500; Santa Cruz Biotechnology), were used as primary antibodies. Horseradish peroxidase-conjugated anti-goat antibody (1:4000; Kierkegaard & Perry Laboratories, Gaithersburg, MD, USA) was used to probe for COX-2 and eNOS, and anti-rabbit antibody (1:2,000; Santa Cruz Biotechnology) was used to probe for iNOS. Band detection was performed using enhanced chemiluminescence detection system (Santa Cruz Biotechnology).
Determination of NO Production
To determine the effect of rocuronium bromide on NO production, the concentration of nitrite in the supernatant was measured using a commercially available NO detection kit (iNtRON Inc., Seoul, Korea), according to the previously described method [14]. After collection of 100 μL of supernatant, 50 μL of N1 buffer was added to each well, and the plate was incubated at room temperature for 10 minutes. Then, N2 buffer was added, and the plate was incubated at room temperature for 10 minutes. The absorbance of the contents of each well was measured at a wavelength of 540 nm. The nitrite concentration was calculated from a nitrite standard curve.
Measurement of Prostaglandin E2 Synthesis
Assessment of PGE2 synthesis was performed using a commercially available PGE2 competitive enzyme immunoassay kit (Amersham Pharmacia Biotechnology Inc., Piscataway, NJ, USA), according to the previously described method [14]. Supernatant (100 μL) from the culture medium and standards were added to different wells of a goat anti-mouse IgG coated microtiter plate provided in the kit. Mouse anti-PGE2 antibody and peroxidase-conjugated PGE2 were added to each well, and the plate was incubated at room temperature with shaking for 1 hour. The wells were drained and washed, and 3,3’,5,5’-tetramethylbenzidine/hydrogen peroxide solution was then added. The plate was incubated at room temperature with shaking, and the reaction was stopped after 30 minutes by adding H2SO4. The absorbance of the contents of each well was then measured at a wavelength of 450 nm.
Statistical Analysis
The results are presented as mean±standard error of mean. The data were analyzed by one-way analysis of variance followed by Duncan post hoc test. The differences were considered statistically significant at P<0.05.
RESULTS
Effect of Rocuronium Bromide on the Viability of CPAE Cells
The cells were cultured with rocuronium bromide at the final concentrations of 1, 5, 10, 50, 100, 500, and 1,000 μg/mL for 24 hours, and MTT assay was then carried out. The cells cultured in rocuronium bromide-free media were used as the control. The viability of cells incubated with rocuronium bromide at concentrations of 1, 5, 10, 50, 100, 500, and 1,000 μg/mL was 102.42% ±2.85%, 100.73% ±1.63%, 102.10% ±1.43%, 99.47% ±1.02%, 84.04% ±12.06%, 70.88% ±1.75%, and 45.18%±0.85% of the control value, respectively (Fig. 1). The present results showed that rocuronium bromide exerted no cytotoxicity until it was at a concentration of 50 μg/mL. However, 100 μg/mL, 500 μg/mL, and 1,000 μg/mL of rocuronium bromide reduced the cell viability. Hence, we used 10 μg/mL, 100 μg/mL, 1,000 μg/mL of rocuronium bromide for the next set of experiments.

The effects of rocuronium bromide treatment on the viability of calf pulmonary artery endothelial (CPAE) cells. A, control group; B, 1-μg/mL rocuronium bromide-treated group; C, 5-μg/mL rocuronium bromide-treated group; D, 10-μg/mL rocuronium bromide-treated group; E, 50-μg/mL rocuronium bromide-treated group; F, 100-μg/mL rocuronium bromidetreated group; G, 500-μg/mL rocuronium bromide-treated group; H, 1,000-μg/mL rocuronium bromide-treated group. The results are presented as mean±standard error of mean. *P<0.05 compared to the control group.
Effect of Rocuronium Bromide on COX-1 Expression
The level of COX-1 in the control cells was set as 1.00. The level of COX-1 was increased to 0.94±0.07, 1.01±0.19, 1.08±0.11, and 1.06±0.02 compared to control, after the treatment with rocuronium bromide at 10-, 100- 1,000-, and 2-μg/mL LPS for 24 hours (Fig. 2). The present results show that rocuronium bromide and LPS treatment exerted no significant effect on COX-1 expression in CPAE cells.
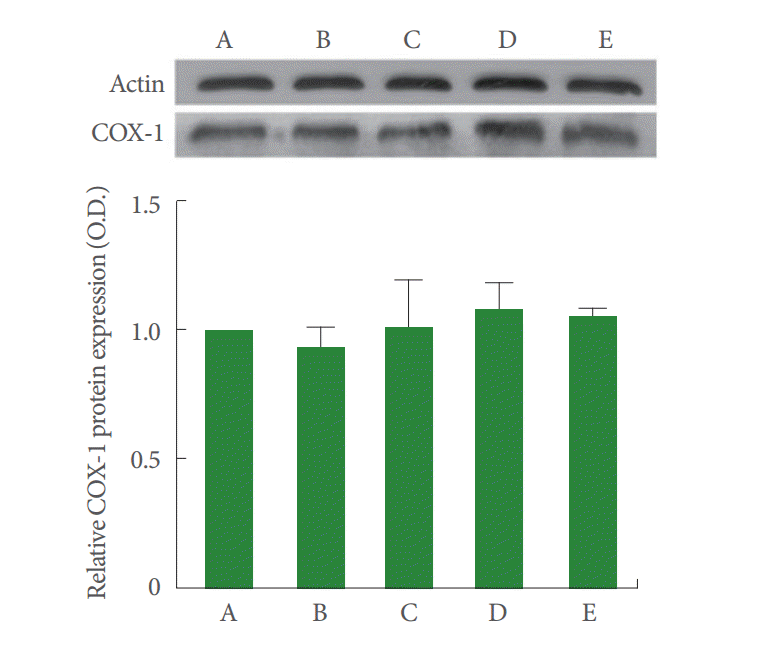
The effects of rocuronium bromide treatment on cyclooxygenase-1 (COX-1) in calf pulmonary artery endothelial (CPAE) cells. A, Control group; B, 10-μg/mL rocuronium bromide-treated group; C, 100-μg/mL rocuronium bromide-treated group; D, 1,000-μg/mL rocuronium bromide-treated group; E, 2-μg/mL lipopolysaccharide-treated group. The results are presented as mean±standard error of mean.
Effect of Rocuronium Bromide on COX-2 Expression
The level of COX-2 in the control cells was set as 1.00. The level of COX-2 was increased to 0.92±0.02, 1.50±0.08, 1.73±0.07, and 1.63±0.11 compared to control after the treatment with rocuronium bromide at 10-, 100- 1,000-, and 2-μg/mL LPS for 24 hours (Fig. 3). The present results show that rocuronium bromide and LPS treatment enhanced COX-2 expression in CPAE cells.
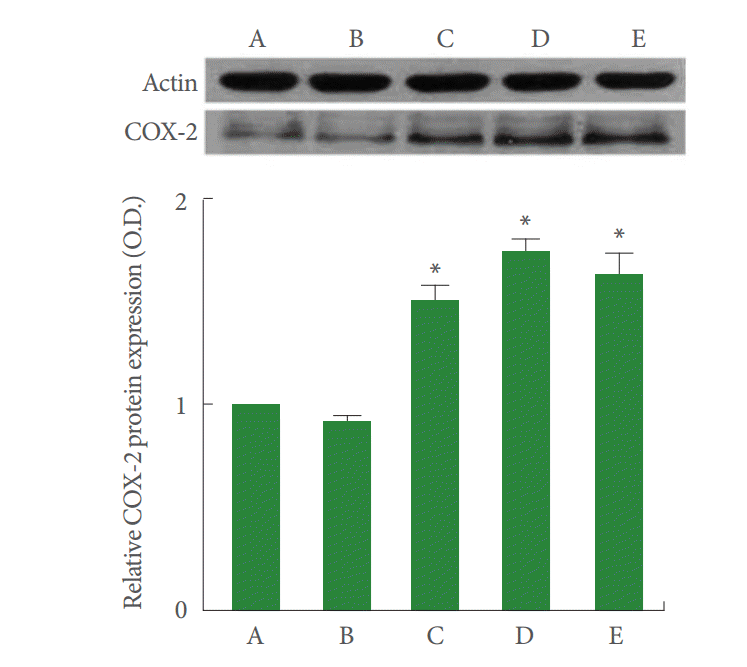
The effects of rocuronium bromide treatment on cyclooxygenase-2 (COX-2) in calf pulmonary artery endothelial (CPAE) cells. A, Control group; B, 10-μg/mL rocuronium bromide-treated group; C, 100-μg/mL rocuronium bromide-treated group; D, 1,000-μg/mL rocuronium bromide-treated group; E, 2-μg/mL lipopolysaccharide-treated group. The results are presented as mean±standard error of mean. *P<0.05 compared to the control group.
Effect of Rocuronium Bromide on iNOS Expression
The level of iNOS in the control cells was set as 1.00. The level of iNOS was increased to 1.23 ±0.18, 1.97 ±0.26, 2.13 ±0.32, and 2.27±0.08 compared to control after the treatment with rocuronium bromide at 10-, 100- 1,000-, and 2-μg/mL LPS for 24 hours (Fig. 4). The present results show that rocuronium bromide treatment enhanced iNOS expression in CPAE cells.

The effects of rocuronium bromide treatment on inducible nitric oxide synthase (iNOS) in calf pulmonary artery endothelial (CPAE) cells. A, Control group; B, 10-μg/mL rocuronium bromide-treated group; C, 100-μg/mL rocuronium bromide-treated group; D, 1,000-μg/mL rocuronium bromidetreated group; E, 2-μg/mL lipopolysaccharide-treated group. The results are presented as mean±standard error of mean. *P<0.05 compared to the control group
Effect of Rocuronium Bromide on eNOS Expression
The level of eNOS in the control cells was set as 1.00. The level of eNOS was increased to 0.88±0.00, 0.63±0.02, 0.26±0.11, and 0.37±0.08 compared to control after the treatment with rocuronium bromide at 10-, 100- 1,000-, and 2-μg/mL LPS for 24 hours (Fig. 5). The present results show that rocuronium bromide and LPS treatment enhanced eNOS expression in CPAE cells.

The effects of rocuronium bromide treatment on endothelial nitric oxide synthase (eNOS) in calf pulmonary artery endothelial (CPAE) cells. A, Control group; B, 10-μg/mL rocuronium bromide-treated group; C, 100-μg/mL rocuronium bromide-treated group; D, 1,000-μg/mL rocuronium bromidetreated group; E, 2-μg/mL lipopolysaccharide-treated group. The results are presented as mean±standard error of mean. *P<0.05 compared to the control group.
Effect of Rocuronium Bromide on NO Production
NO production was decreased to 2.91±0.08, 2.96±0.16, 2.46±0.05, and 1.96±0.07 μM after rocuronium bromide treatment at 10-, 100-, 1,000-, and 2-μg/mL LPS for 24 hours. In contrast, LPS increased NO production to 3.71±0.06 μM (Fig. 6). The present results showed that rocuronium bromide treatment decreased NO production, while LPS treatment increased NO production in CPAE cells.

Measurement of nitric oxide (NO) production in calf pulmonary artery endothelial (CPAE) cells. A, Control group; B, 10-μg/mL rocuronium bromide-treated group; C, 100-μg/mL rocuronium bromide-treated group; D, 1,000-μg/mL rocuronium bromide-treated group; E, 2-μg/mL lipopolysaccharidetreated group. The results are presented as mean±standard error of mean. *P<0.05 compared to the control group.
Effect of Rocuronium Bromide on PGE2 Synthesis
PGE2 synthesis was increased to 55.97 ±0.91, 59.07 ±2.26, 64.17±0.86, 71.29±1.10, and 96.73±1.41 pg/well after the treatment with rocuronium bromide at 10-, 100- 1,000-, and 2-μg/mL LPS for 24 hours (Fig. 7). The present results showed that rocuronium bromide and LPS treatment increased PGE2 synthesis in CPAE cells.

Measurement of prostaglandin E2 (PGE2) synthesis in calf pulmonary artery endothelial (CPAE) cells. A, Control group; B, 10-μg/mL rocuronium bromide-treated group; C, 100-μg/mL rocuronium bromide-treated group; D, 1,000-μg/mL rocuronium bromide-treated group; E, 2-μg/mL lipopolysaccharidetreated group. The results are presented as mean±standard error of mean. *P<0.05 compared to the control group.
DISCUSSION
Acidic solutions were suggested as the underlying mechanism of rocuronium bromide-induced injection pain [4]. However, Borgeat and Kwiatkowski [1] reported that low pH is unlikely to be the cause of injection pain because patients receiving normal saline buffered to pH 4 did not complain about pain. Local mediators, such as kinins that directly irritate the venous nociceptors and the allogenic effect of aminosteroid neuromuscular blocking drugs were suggested as the possible mechanisms of rocuronium bromide-induced injection pain [16]. Park et al. [17] demonstrated that remifentanil effectively prevents rocuronium-induced injection pain or withdrawal movement. Zhang et al. [18] reported that the incidence and severity of rocuronium-induced injection pain were significantly alleviated by use of a large vein. However, most commonly investigated pharmacological interventions to reduce rocuronium-induced injection pain are of low quality due to risk of bias and inconsistency.
Endothelial cells are known to possess both isoforms of COX, and their induction has been demonstrated to occur in response to different proinflammatory cytokines, such as interleukin (IL)-1α and IL-β and tumor necrosis factor-α [19]. COX-2 is induced by inflammatory stimuli and synthesizes prostaglandins, which modulate vascular tone and mediate the inflammatory process and/or tissue damage. Specific COX-2 inhibitors attenuate the symptoms of inflammation [6]. In the present study, rocuronium bromide and LPS treatment exerted no significant effect on COX-1 expression; in contrast, rocuronium bromide and LPS treatment increased COX-2 expression in CPAE cells. These results suggest that rocuronium bromide might initiate inflammation through COX-2 activation in the vascular wall.
iNOS is involved in hyperalgesia and neuropathic pain [20]. iNOS induction plays an integral role in mediating endothelial dysfunction [21]. In the present study, rocuronium bromide and LPS treatment increased iNOS expression in CPAE cells. These results suggest that rocuronium bromide might induce hyperalgesia state in the vascular wall.
Replicative aging results in decreased expression of eNOS, which is associated with endothelial dysfunction and increased risk for atherosclerosis [22]. eNOS down-regulation increased expression of leukocyte adhesion molecules, resulting in inflammation [22,23]. Weak eNOS immunoreactivity in the endothelium causes structural alteration of the vascular wall and vasoconstriction [24]. Simultaneous down-regulation of eNOS and up-regulation of iNOS may increase the inflammatory response of endothelial cells [25]. In the present study, rocuronium bromide and LPS treatment decreased eNOS expressions in CPAE cells. These results suggest that rocuronium bromide might accelerate inflammatory conditions in the vascular wall.
NO is a potent vasodilator that regulates vascular tone, and NO potentially exerts vasoprotective effect in the vascular wall [10,11]. Reduced NO production is related to microcirculation disturbance [26,27]. In contrast, high concentrations of NO have a detrimental effect on the inflammation microenvironment [28]. This biphasic aspect of cytoprotective or cytotoxic effect of NO is driven due to the different production mechanisms. In the present study, rocuronium bromide treatment decreased NO synthesis in CPAE cells. On the other hand, LPS treatment increased NO synthesis. Reduction in NO synthesis may result from reduced expression of eNOS in CPAE cells. In the present study, rocuronium bromide treatment inhibited NO production, while LPS treatment increased NO production in CPAE cells. These results suggest that decreased NO production after rocuronium treatment might cause stasis of rocuronium bromide in the vascular wall.
PGE2 is implicated in the pathogenesis of pain in acute and chronic inflammatory disease states [29]. Production of PGE2 causes pain and suppression of PGE2 is related with analgesic effect [15]. In the present study, rocuronium bromide and LPS increased PGE2 production in CPAE cells. These results suggest that rocuronium bromide treatment might produce pain in the vascular wall.
In the present results, rocuronium bromide inhibited eNOS, and then suppressed NO production in CPAE cells. Rocuronium bromide activated COX-2, iNOS, and then increased PGE2 synthesis in CPAE cells. There is a time difference between the present results and rocuronium bromide-induced injection pain or withdrawal movement. However, rocuronium bromide might induce inflammation and pain in CPAE cells. Suppressing NO production and enhancing PGE2 synthesis might be associated with rocuronium bromide-induced injection pain or withdrawal movement.
Notes
Conflict of Interest
No potential conflict of interest relevant to this article was reported.