Altered Secretory Activity of APE1/Ref-1 D148E Variants Identified in Human Patients With Bladder Cancer
Article information

Abstract
Purpose:
Apurinic/apyrimidinic endonuclease 1/redox factor-1 (APE1/Ref-1) is a multifunctional protein involved in DNA repair and redox modulation. Recently, serum and urinary APE1/Ref-1 levels were reported to be increased in patients with bladder cancer. Genetic variations of APE/Ref-1 are associated with the risk of cancer. However, the effect of APE1/Ref-1 variants on its secretory activity is yet unknown.
Methods:
APE1/Ref-1 variants were evaluated by DNA sequencing analysis of reverse transcription polymerase chain reaction products in coding DNA sequences (CDS) of APE1/Ref-1 in bladder tissue samples from patients with bladder cancer (n=10). Secretory activity of APE1/Ref-1 variants was evaluated with immunoblot and enzyme-linked immunosorbent assay of the culture medium supernatants.
Results:
Four different substitution mutants (D148E, I64V/D148E, W67R/D148E, and E86G/D148E) of APE1/Ref-1 were identified in bladder cancer specimens. However, deletion mutants of APE1/Ref-1 CDS were not found. The secretory activity of the APE1/Ref-1 variants (D148E, I64V/D148E, and E86G/D148E) was increased compared to that of wild type APE1/Ref-1. Furthermore, the secretory activity in basal or hyperacetylated conditions was much higher than that in APE1/Ref-1 D148E-transfected HEK293 cells.
Conclusions:
Taken together, our data suggest that the increased secretory activity of D148E might contribute to increased serum levels of APE1/Ref-1 in patients with bladder cancer.
INTRODUCTION
Bladder cancer is a common urologic cancer prevailing in Korea [1]. Early detection of bladder cancer is critical to improve the survival rate. Although cystoscopy is the gold standard for detection of bladder cancer, the development of noninvasive serologic or urinary biomarkers would greatly aid in the diagnosis or treatment of bladder cancer.
The human apyrimidinic endonuclease 1/redox factor-1 (APE1/Ref-1) gene is located on chromosome 14q11-12, and encodes a 318 aa long protein with a molecular weight of 36 kDa [2]. APE1/Ref-1 plays an important role in DNA repair and redox regulation. The C-terminus of APE1/Ref-1 is involved in DNA repair as an apurinic/apyrimidinic endonuclease. In contrast, the N-terminus is involved in redox regulation as a reducing donor to cysteine residues, leading to the modulation of transcriptional factors, such as activator protein-1 and nuclear factor-kappa B, and antioxidant activity [3]. Oxidative stress, tobacco exposure, and/or environmental chemicals can induce DNA damage, leading to tumorigenesis in human tissues. APE1/Ref-1 plays an important role against oxidative or environmental DNA damage. APE1/Ref-1 genetic variation is closely linked to cancer pathogenesis or susceptibility [4]. Single nucleotide polymorphism analyses show that APE1/Ref-1 polymorphisms may contribute to an increased risk of bladder cancer [5,6].
The APE1/Ref-1 protein level is increased in the serum and urine of patients with bladder cancer [7,8], which can be useful for the diagnosis of bladder cancer. Increased APE1/Ref-1 levels are associated with the tumor stage and grade of bladder cancer, suggestive of being a reliable biomarker of bladder cancer [7,8]. The precise mechanism underlying the existence of serum or urinary APE1/Ref-1 in patients with bladder cancer is still unknown. However, secretion of APE1/Ref-1 is closely related with intracellular acetylation [9-11].
Therefore, we aimed to investigate and identify APE1/Ref-1 gene variants in patients with bladder cancer. The effect of APE1/Ref-1 variants on the secretory activity of APE1/Ref-1 was investigated in the supernatant of cultured cells.
MATERIALS AND METHODS
Reagents
Trichostatin A (TSA), Ponceau S, 4ʹ,6-diamidino-2-phenylindole (DAPI), and fluorescence mounting solution were purchased from Sigma-Aldrich (St. Louis, MO, USA). Dulbecco’s modified Eagle’s medium (DMEM) was purchased from Welgene (Seoul, Korea). Fetal bovine serum, antibiotics, Opti-MEM, methanol-free paraformaldehyde, and transfection reagent, Lipofectamine LTX with Plus were purchased from Thermo Fisher Scientific (Waltham, MA, USA). Antibodies against FLAG and β-actin were purchased from Sigma-Aldrich. Two different APE1/Ref-1 antibodies were used. A monoclonal antibody for the detection of APE1/Ref-1 (N-terminal 80–100 aa) was obtained from Novus (Littleton, CO, USA). A polyclonal rabbit APE1/Ref-1 antibody was generated by immunization with recombinant human APE1/Ref-1 (rh APE1/Ref-1) as previously reported [11].
Identification of APE1/Ref-1 Variants in Bladder Cancer Tissues
Bladder tissue biospecimens were obtained from patients with operable bladder cancer (n =10). All samples were obtained from the archives of the Department of Urology, Chungnam National University, between 2010 and 2011. This study was approved by the Institutional Review Board of Chungnam National University Hospital (approval number: 1101-16). To identify human APE1/Ref-1 variants in coding DNA sequences, reverse transcription polymerase chain reaction was performed; Polymerase chain reaction (PCR) primers were designed based on the GenBank reference sequences (accession number. NM_001641). The coding DNA sequence of human APE1/Ref-1 cDNA was amplified using the following primers set: forward primer 5ʹ-TGC CCA AGG GCG TTC GTA ACG GGA -3ʹ, reverse primer 5ʹ-GCT CAA AGT GAT TTA GGG GTG GTG -3ʹ. Amplified cDNA was inserted into the pGEM-T easy vector system (Promega, Madison, WI, USA) and transformed into DH5α Escherichia coli. Finally, APE1/Ref-1 variants were identified with DNA sequencing analysis. Furthermore, to evaluate the function of APE1/Ref-1 variants, the identified human APE1/Ref-1 variants were subcloned into a pCMV-Tag2A vector between EcoRI and HindIII restriction sites or a pEGFP-C1 vector between EcoRI and KpnI restriction sites. The subcloning of each cDNA was confirmed by DNA sequencing.
Cell Culture and Transfection
The human embryonic kidney epithelial 293T (HEK293T) cell line was purchased from the American Type Culture Collection (Manassas, VA, USA). Cells were grown in DMEM (Welgene) containing 10% fetal bovine serum (Thermo Fisher Scientific) and 1% antibiotics. Cells were maintained in a humidified atmosphere of 95% air and 5% CO2, at 37°C. For transient transfection, HEK293T cells were seeded in plates at a density of 2.5×105 cells/mL and incubated overnight at 37°C. HEK293T cells were transfected with each APE1/Ref-1 construct using Lipofectamine LTX (Thermo Fisher Scientific) according to the manufacturer’s protocol.
Determination of Cell Viability
Cell viability was determined using propidium iodide staining and an automatic cell counter (ADAM-MC), as previously described [9].
Measurement of Secreted APE1/Ref-1
A sandwich enzyme-linked immunosorbent assay (ELISA) was used to quantify the level of secreted APE1/Ref-1 in the culture medium (CM), as previously described [11].
Immunoprecipitation
HEK293T cells were grown to confluence on a 6-well plate (5×105 cells/well) and then treated with 1μM TSA for 1 hour after changing the CM. The CM was collected at different time points and centrifuged at 1,200 rpm for 3 minutes to remove the residual cells. The supernatant was again centrifuged at 3,000 rpm for 3 minutes. The APE1/Ref-1 in the CM was immunoprecipitated using anti-APE1/Ref-1 antibody as previously reported [10]. The immune complexes were mixed with sample buffer and subjected to sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) followed by immunoblotting.
Immunoblotting
Whole cell lysates were prepared as previously described [11]. The cell lysates were cleared by centrifugation at 14,000×g for 15 minutes, and the supernatant fraction was used for immunoblotting. Proteins were resolved by SDS-PAGE and transferred onto a polyvinylidene difluoride membrane. After blocking with 5% nonfat dry milk in Tris-buffered saline (TBS) containing 0.05% Tween 20, the membrane was incubated with the desired primary antibody overnight at 4°C. The membrane was then washed and probed with the appropriate secondary antibody, and the immunoreactive bands were visualized by an enhanced chemiluminescence method. Each membrane was stripped and reprobed with antiactin antibody to ensure equal protein loading.
Subcellular Localization of APE1/Ref-1 Variants
A subcellular localization of each APE1/Ref-1 variant was observed in cells transfected with EGFP-tagged APE1/Ref-1 variants as previously described [12]. Cells were grown on glass coverslips and transiently transfected with 1.6 μg of plasmid from the pEGFP-tagged APE1/Ref-1 variant plasmids. Cells were fixed with 4% paraformaldehyde, and DAPI was used for 10 minutes to stain nucleus. Coverslips were mounted on microscopic slides using a fluorescence mounting solution and green fluorescence protein (GFP) signals were visualized with a fluorescence microscope (Carl Zeiss, Jena, Germany).
Statistical Analysis
All data are expressed as the mean±standard error of the mean. Statistical significance of differences in measured variables was determined using one-way analysis of variance followed by Bonferroni test for multiple comparisons. Differences were considered significant at P<0.05.
RESULTS
Identification of APE1/Ref-1 Gene Variants in Patients With Bladder Cancer
Nucleotide variants of APE1/Ref-1 cDNA were evaluated in 10 patients with bladder cancer. DNA sequencing analysis data showed the identification of four different APE1/Ref-1 variants: a single mutation with Asp/Glu at codon 148 of APE1/Ref-1 (D148E), and 3 different double mutations with Ile/Val at codon 64 (I64V), Trp/Arg at codon 67 (W67R), and Glu/Gly at codon 86 (E86G) with D148E (Table 1). Furthermore, APE1/Ref-1 variants with nucleotide substitutions 318 T>A or 257 A>G were observed without amino acid changes in 2 specimens. Noticeably, the genotype with D148E was observed in 6 specimens with single or double mutations (Table 1).
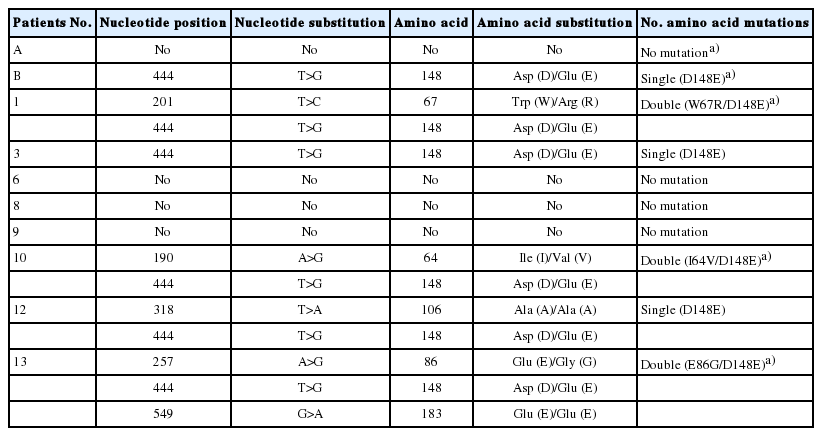
Nucleotide and amino acid mutations identified in the coding DNA sequence of APE1/Ref-1 of patients with human bladder cancer
Protein Expression and Localization of APE1/Ref-1 Variants
To examine the association of APE1/Ref-1 variants in relation to its expression in cells, we constructed four expression vectors containing full APE1/Ref-1 cDNA with D148E, I64V/D148E, W67R/D148E, and E86G/D148E. The level of APE1/Ref-1 protein expression was determined in 293T cells, which were transiently transfected with the substitution mutant constructs. As shown in Fig. 1A, the APE1/Ref-1 proteins were observed without any fragmentation in cells expressing each substitution mutant. Lower APE1/Ref-1 protein expression compared to substitution mutants was only detected in W67R/D148E expressing cells, although the differences in protein levels were normalized against the actin loading control.
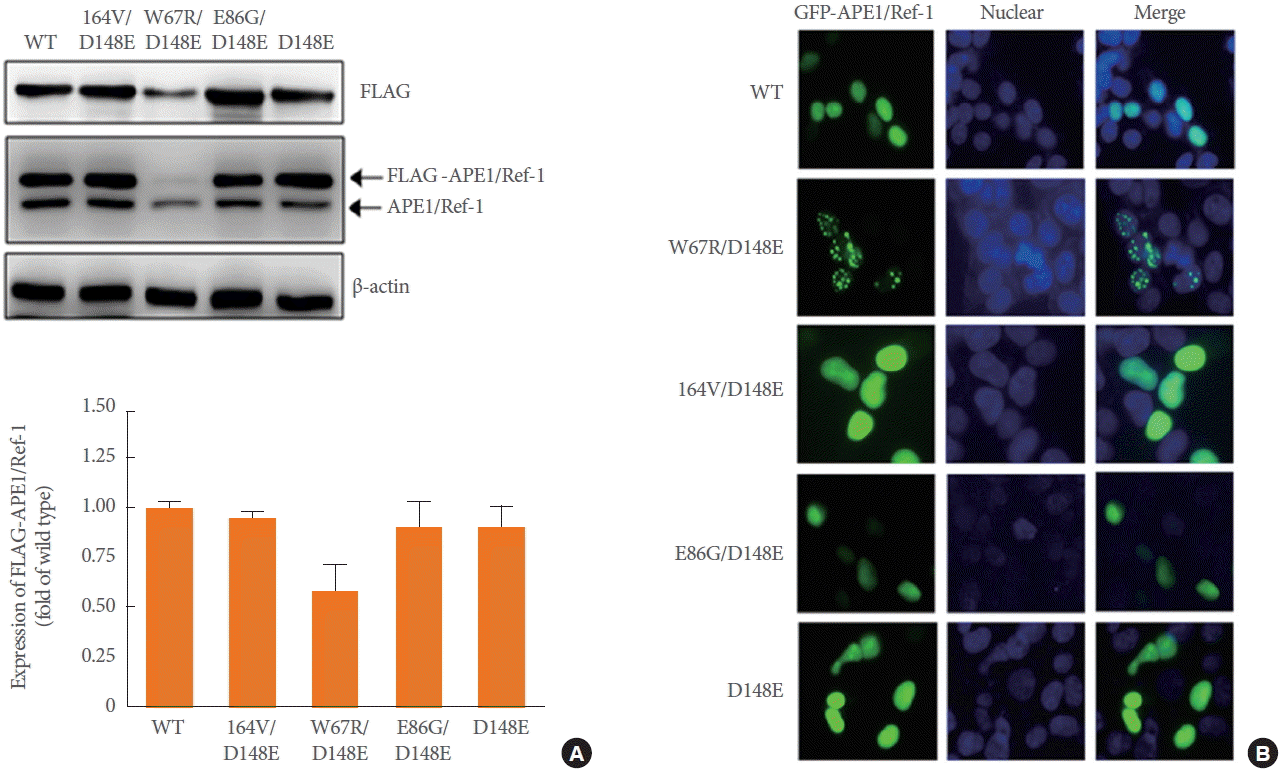
Expression of APE1/Ref-1 variants in HEK293T cells transiently transfected with FLAG tagged-wild-type (WT) or one of 4 constructs: W67R/D148E, I64V/D148E, E86G/D148E, or D148E. (A) Immunoblot analysis using anti-FLAG or anti-APE1/Ref-1 antibody was performed. Anti-β-actin was used as loading control. Similar results were observed in experiments run in triplicate. (B) Intracellular localization of GFP-APE1/Ref-1 variants in HEK293T cells. HEK293T cells were transfected with GFP-tagged (green) WT or one of 4 constructs. Cells were fixed with 4% paraformaldehyde and the nuclei were stained with DAPI (blue) (×400). Similar results were observed in experiments run in duplicate. APE1/Ref-1, apurinic/apyrimidinic endonuclease 1/redox factor-1; HEK293T, human embryonic kidney epithelial 293T; GFP, green fluorescent protein; DAPI, 4ʹ,6-diamidino-2-phenylindole.
The effect of APE1/Ref-1 variants on cellular localization was further investigated using GFP-tagged substitution mutant constructs. Representative images for GFP-APE1/Ref-1, and DAPI staining in 293T cells expressing each substitution mutant was shown in Fig. 1B. Consistent with immunoblot results, each APE1/Ref-1 mutant exclusively existed in the nuclei as evident from the colocalization of green (APE1/Ref-1 variants) and blue fluorescence (DAPI staining). Remarkably, the W67R/D148E double mutant was distributed as random-dots in the nucleus, indicating a possible alteration in the localization of APE1/Ref-1 caused by the substitution mutation.
Extracellular Secretion of APE1/Ref-1 Variants
We proposed earlier that the secretory APE1/Ref-1 is a novel biomarker because the level of APE1/Ref-1 protein is elevated in the serum and urine samples of patients with bladder cancer [7,8]. Indeed, many reports have corroborated the extracellular existence and dynamic translocalization of APE1/Ref-1 from the nuclei to cytosol [11,12]. Here, we determined whether the substitution mutant of APE1/Ref-1 was secreted into CM. The secretion level of each APE1/Ref-1 mutant in CM was estimated by comparison with a recombinant APE1/Ref-1 protein standard curve. As shown in Fig. 2A, the cell viability in basal or TSA-stimulated conditions was not changed in 293T cells transfected with APE1/Ref-1 variants. The secretory activity of APE1/Ref-1 variants was evaluated using the newly developed ELISA [7] in basal and TSA-stimulated conditions. Basal secretion of APE1/Ref-1 in cells transfected with substitution mutants (I64V/D148E, E86G/D148E, and D148E) was significantly increased compared to that of wild-type APE1/Ref-1 (Fig. 2B). We previously reported that the acetylation of cellular proteins by histone deacetylase (HDAC) inhibitors, TSA, and acetylating agents induced the extracellular release of APE1/Ref-1 [9]. Interestingly, all variants with D148E, except double mutant W67R/D148E, showed increased APE1/Ref-1 secretion in response to TSA.
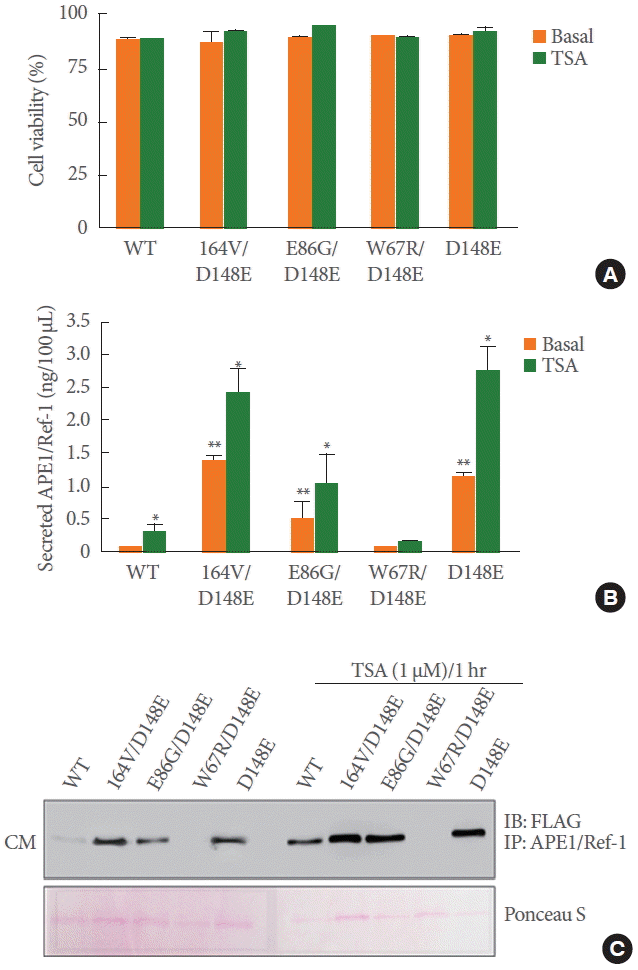
Extracellular secretion of APE1/Ref-1 variants from HEK293T cells transiently transfected with FLAG tagged-wild-type (WT) or one of 4 constructs: W67R/D148E, I64V/D148E, E86G/D148E, or D148E. HEK293T cells expressing WT or variant APE1/Ref-1 treated with 1μM TSA for 1 hour. (A) Effect of TSA on cell viability was determined using an automatic cell counter. (B) The CM was collected without cell debris and the level of secreted APE1/Ref-1 was measured by ELISA. Columns, mean (n=3); bars, standard error. *P<0.05, significantly different compared to TSA nontreated cells; **P<0.01 significantly different compared to WT transfected cells by one-way analysis of variance followed by Bonferroni multiple comparison test. (C) The collected CM was immunoprecipitated with anti-APE1/Ref-1 followed by monoclonal anti-FLAG antibodies. Ponceau S was used as the loading control. APE1/Ref-1, apurinic/apyrimidinic endonuclease 1/redox factor-1; HEK293T, human embryonic kidney epithelial 293T; TSA, trichostatin A; CM, culture medium; ELISA, enzyme-linked immunosorbent assay; DAPI, 4',6-diamidino-2-phenylindole.
Immunoprecipitation using specific anti-APE1/Ref-1 antibody followed by immunoblot was performed to visualize the secreted APE1/Ref-1 in the CM. Minimal secreted APE1/Ref-1 was detected in native culture supernatants including wild-type APE1/Ref-1. The maximum secretion level of APE1/Ref-1 variants with the D148E mutation, although it was in its nonacetylated form. The expression of secreted APE1/Ref-1 in all variants was remarkably increased with exposure of cells to TSA treatment. However, the secretion of APE1/Ref-1 in double mutant W67R/D148E was not measured. Taken together, these data indicate that APE1/Ref-1 secretion is obviously caused by mutation at D148E of APE1/Ref-1, implying a correlation between D148E mutation with increased secretory activity and its role as a biomarker for bladder cancer.
Kinetics of Secreted APE1/Ref-1 D148E Variants
Given that the secretion of D148E in APE1/Ref-1 was clearly distinct from other secretions, we directly compared the secreted amount of wild-type and D148E variants in CM for 1 hour after changing media, as represented in Fig. 3A (upper). The secretion level of the wild-type APE1/Ref-1 protein reached 0.15 ng/mL in 1 hour, whereas that of the D148E mutants showed a drastic increase to 0.65 ng/mL in the same amount of time, suggesting a high secretory activity of D148E mutants even in the absence of any stimulus (Fig. 3A bottom). Next, degradation kinetics of secreted APE1/Ref-1 D148E mutants were studied and evaluated for 6 hours in cell-free medium after stimulation with TSA for 1 hour. Nonlinear fit regression analysis showed that the half-life for degradation of the secreted wild-type and D148E was not significantly different (half-life of 1.6 hours for wild-type and 1.8 hours for D148E) in cell-free CM (Fig. 3B). Collectively, these results suggest that the increased levels of APE1/Ref-1 in the D148E variant might be due to increased secretory activity, rather than the delayed degradation of D148E.
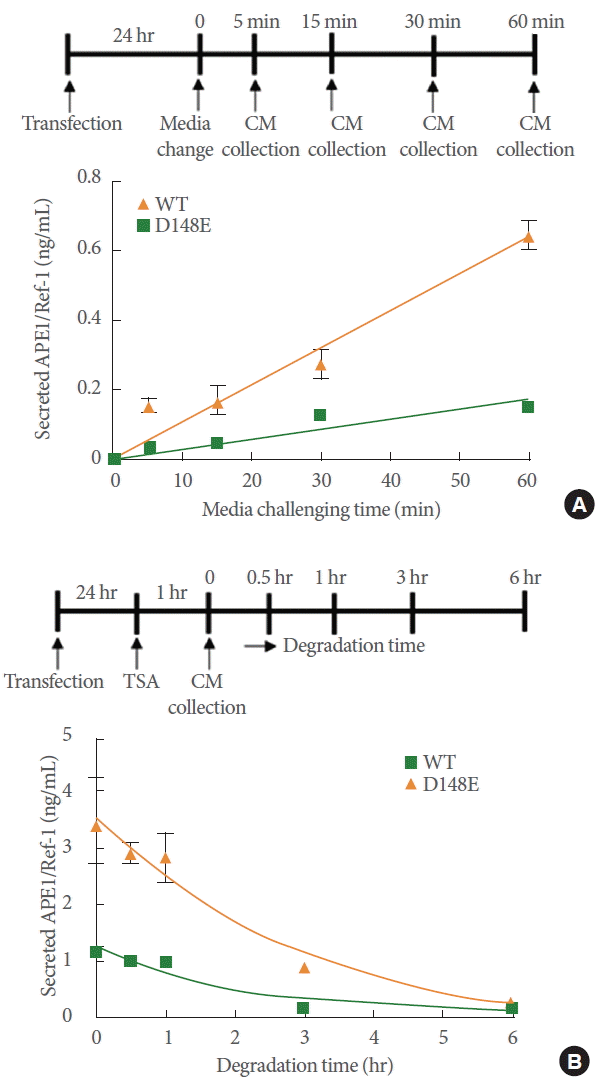
Comparison of secretion activity and stability of wild-type (WT) APE1/Ref-1 and the D148E variant. (A) HEK293T cells expressing WT or D148E variant APE1/Ref-1 were prepared. After transfection for 24 hours, the CM was replaced with fresh media and then time-dependently collected as shown in the scheme. (B) HEK293T cells expressing WT or D148E variant APE1/Ref-1 were treated with 1 μM TSA for 1 hour. The CM was transferred into an empty plate and 0.5 mL of this CM was time-dependently collected for 6 hours as shown in the scheme. The level of APE1/Ref-1 in the CM was measured by ELISA. These experiments were performed in triplicate with similar results. APE1/Ref-1, apurinic/apyrimidinic endonuclease 1/redox factor-1; HEK293T, human embryonic kidney epithelial 293T; CM, culture medium; TSA, trichostatin A; ELISA, enzyme-linked immunosorbent assay.
DISCUSSION
In the present study, we identified 4 distinct APE1/Ref-1 variants: one with a single substitution mutation in D148E and 3 with double substitution mutations in I64V/D148E, W67R/D148E, and E86G/D148E in bladder tissue specimens from patients with bladder cancer. Using DNA recombination technology, all substitution mutations identified in bladder cancer were subcloned, and their functional role on the secretion of APE1/Ref-1 was evaluated.
Risk factors such as cigarette smoking and chronic exposure to toxic environmental chemicals play key roles in the tumorigenesis of bladder cancer [13]. DNA damage on single-strand breaks is mainly repaired with base excision repair proteins like APE1/Ref-1 [14]. Genetic variation of APE1/Ref-1 is associated with the risk of cancer. Among the genetic variations, the single substitution D148E has been extensively investigated in association with different types of cancer, such as lung, breast, gastric, and bladder cancers [5,15-17].
In the present study, the most common mutation of APE1/Ref-1 was identified as D148E, which is found in approximately 60% of patients with bladder cancer. However, double mutations of APE1/Ref-1 were also identified at positions I64V, W67R, and E86G. Protein expression of three substitution mutants of APE1/Ref-1 (D148E, I64V/D148E, and E86G/D148E) was found to be similar to that of wild-type APE1/Ref-1. However, the protein expression in the event of the double mutation of APE1/Ref-1 (W67R/D148E) was dramatically decreased, suggesting an inhibitory role of W67R in APE1/Ref-1 protein expression. Subcellular localization of all four substitution mutants of APE1/Ref-1 (D148E, I64V/D148E, E86G/D148E, and W67R/D148E) was mainly confined to the nucleus, similar to that of wild-type APE1/Ref-1. However, the subcellular localization of APE1/Ref-1 (W67R/D148E) was observed as condensed forms in the nucleolus of nuclei. Mutations in the putative nuclear localization signals in the N-terminal 20-residue portion [18] did not occur in the four substitution mutants of patients with bladder cancer. However, the W67R mutation, a mutation of region containing nuclear export signals (64-80 aa of APE1/Ref-1) [19], may affect subcellular localization and intracellular trafficking of APE1/Ref-1.
We also evaluated the effect of mutations on APE1/Ref-1 secretion. APE1/Ref-1 can be secreted in the CM in response to intracellular hyperacetylation, such as with aspirin or HDAC inhibitors [9,10]. The expression of APE1/Ref-1 is enhanced in the serum and urine of patients with bladder cancer [7,8]. Interestingly, the secretory activity of D148E mutant APE1/Ref-1 is significantly increased compared to that of wild-type APE1/Ref-1. Although the precise mechanism behind the increased secretory activity of APE1/Ref-1 D148E is not yet known, there are some hypotheses regarding the subject. In silico analyses indicate that the Asp148Glu substitution may cause changes in the protein structure and is associated with an increased risk of breast cancer [20]. The target amino acid at position 148 of the APE1/Ref-1 protein was mutated from Asp to Glu and selected for the lowest energy conformations [20]. The D148E polymorphism of APE1/Ref-1 is related to reduced base excision repair efficiency, which is potentially linked with genomic instability [21]. In addition, the amino acid substitutions may alter the redox function of APE1/Ref-1. Therefore, this genomic instability and/or alteration of redox function, such as changes in transcriptional activity, can influence the secretory activity of D148E.
There is dynamic crosstalk between APE1/Ref-1 and the intracellular acetylation system. APE1/Ref-1 is acetylated by histone acetyl transferase of p300 [22] and is deacetylated by Sirtuin-1, a protein deacetylase, associated with APE1/Ref-1 [23]. Moreover, APE1/Ref-1 can induce Sirtuin-1 activation via redox [24,25]. Therefore, genetic variations in APE1/Ref-1 can influence intracellular acetylation status [9]. TSA is a well-known HDAC inhibitor and is used to induce the secretion of APE1/Ref-1 in cultured cell systems [9]. In the present study, we successfully demonstrated that the secretory activity of APE1/Ref-1 D148E is greater than that of wild-type APE1/Ref-1, with basal acetylation levels in response to TSA, suggesting that the increased secretion of APE1/Ref-1 D148E may be due to increased sensitivity of intracellular acetylation by HDAC inhibitors. Our findings in this study regarding the secretory activity of APE1/Ref-1 D148E are important, as they may aid in understanding the potential role of fundamental genetic variants in the increased serologic APE1/Ref-1 levels observed in certain disorders like malignant tumors.
Notes
Grant Support
This research was supported by the Basic Science Research Program through the National Research Foundation of Korea (NRF-2014R1A6A1029617).
Research Ethics
This study was approved by the Institutional Review Board of Chungnam National University Hospital (approval number: 1101-16). The biospecimens and data used for this study were provided by the Biobank of Chungnam University Hospital, a member of the Korea Biobank Network.
Conflict of Interest
No potential conflict of interest relevant to this article was reported.