Can Urinary Bladder Innervation Be Restored After Outlet Obstruction and Denervation?
Article information

Abstract
Transurethral resection of the prostate, or other methods to decrease outlet resistance usually leads to relief of symptoms in patients with bladder outlet obstruction (BOO). If symptoms of underactivity persist after normalization of outflow conditions, treatment options are limited. In this review, we hypothesize, based on results from basic research, what might become treatment options for such patients in the future. The primary local treatment will still aim at reducing outlet obstruction. We speculate that local secondary treatment in the future might include transplantation of stem cells or mature bladder ganglion cells into the bladder wall. There has been some success in transplanting ganglion cells into the rat bladder. The ganglion cells will sprout into the surrounding tissue but functional connections between the axons of the transplanted neurons, and the detrusor smooth muscle have so far not been demonstrated. Neurotrophins or neurotrimin might be injected into the bladder wall to increase the sprouting of existing or transplanted neurons. Stem cell transplantation has been performed and improves detrusor function, but it has so far, been difficult to demonstrate transplanted stem cells. BOO, persisting detrusor underactivity, and decreased nerve density are often combined with inflammatory activity of the lower urinary tract. NLR family pyrin domain containing 3 (NLRP3) and its messenger RNA (mRNA) as well as cyclooxygenase-2 (Cox-2) mRNA are increased in obstructed bladders. Systemic treatment with the NLRP3 inhibitor glyburide normalized nerve density in rat bladder, and, to some extent, bladder function. It is unclear whether Cox-2 is involved in the decreased nerve density following obstruction, but Cox-2 mRNA increases 5-fold in obstructed bladder. Future therapy against bladder underactivity remaining following relief of obstruction includes either systemic treatment, perhaps by anti-inflammatory drugs, or local treatment by injection of stem cells, mature ganglion cells, and/or neurotrophins or neurotrimin into the bladder wall.
INTRODUCTION
The motor innervation of the human urinary bladder is mainly by postganglionic parasympathetic axons. The ganglion cells are situated in ganglia both along the parasympathetic nerves and in the bladder itself (intramural ganglia). Such intramural ganglion cells are also found in e.g., guinea pig bladders [1]. The sensory input to the spinal cord is via afferent axons with their perikarya situated in the dorsal root ganglia. The adrenergic innervation is sparse except in the bladder neck and urethral sphincter. The parasympathetic innervation of the bladder is very dense, each detrusor muscle cell is reached by at least one nerve terminal [2]. The innervation of the human bladder is a resilient system. Lesions will affect a mixture of preganglionic (decentralization) and postganglionic (postganglionic denervation) axons, and probably the intramural ganglion cells. Nerve damage of clinical importance might occur e.g., after pelvic organ surgery, pelvic fractures, bladder outlet obstruction, diabetes mellitus, and acrylamide intoxication. In the bladders subjected to outlet obstruction, the lesions are mostly postganglionic but might also affect the intramural ganglion cells. Injury of the afferent nerves might also contribute to bladder dysfunction and is a major cause of bladder dysfunction in diabetes mellitus and acrylamide intoxication.
Denervated and obstructed bladders have similarities. The detrusor muscle cells hypertrophy in both, and the bladder weight increases. Array data show that the hypertrophic response to denervation and obstruction are quite similar. One example is that the messenger RNA (mRNA) for the gap junction protein connexin 43 increases in both (by 2.63 in 10-day obstructed bladder, and by 3.30 in 10-day denervated bladder) [3]. An increased number of connexons between the smooth muscle cells might be a compensatory mechanism counteracting the decrease or absence of motor nerve terminals. A consequence of this is a higher amplitude of the spontaneous contractions in the bladder wall during filling. This overactivity is most pronounced in the denervated bladder, where the tetrodotoxin-resistant phasic contractions can reach almost 60% of the maximal contractile amplitude [4]. Another example of a similar hypertrophic response in the denervated and obstructed bladder is the significantly increased mRNA for (basal lamina) collagen 4 (10 days obstructed 2.9, 10 days denervated 2.8) and elastin (10 days obstructed 4.1, 10 days denervated 6.5) in the obstructed and denervated bladders. In our arrays, there was no difference in mRNA levels for muscarinic receptors in denervated, and obstructed bladders, and their respective controls. Others have, however, found an increased number of muscarinic receptors in obstructed [5] and denervated bladders [6]. In the latter study, the increase was partly due to an upregulation of the M2 receptor [7].
It may be questioned if the hypertrophy of the denervated bladders is due to the denervation as such, or if it is secondary to the stretch of the smooth muscle cells caused by the distension of the paralyzed bladder. Nilvebrant et al. [6] found that bladder denervation led to increased bladder weight and increased number of muscarinic receptors. If the urinary flow was diverted to avoid bladder distension, bladder weight, and muscarinic receptor density remained normal. Thus, the hypertrophic response and the increased density of M receptors were secondary to the stretch of the bladder wall, and not a direct effect of the denervation.
The rat bladder also hypertrophies in response to diabetes mellitus. Whether this hypertrophy is similar to that of denervated or obstructed bladders is somewhat unclear. Earlier studies have interpreted diabetes hypertrophy as a response to the increased functional demand (increased diuresis) of the diabetic bladder [8]. It has recently been suggested that the hypertrophy is a direct effect of the diabetic state on the detrusor smooth muscle cells [9]. Bladder arrays suggest differences between bladder hypertrophy caused by diabetes and polyuria caused by sucrose feeding [10]. Hindi et al. [10] found e.g., a sevenfold increase in brain-derived neurotrophic factor (BDNF) mRNA in diabetic bladders but no effect by sucrose feeding. We have previously reported a 2-fold increase of BDNF mRNA in obstructed bladders [3]. BDNF stimulates sprouting of parasympathetic axons from the pelvic ganglia [11]. Taken together, this might indicate denervation and reinnervation processes in both diabetic and obstructed bladders. This is supported by the observations that neurotrimin mRNA is increased both in 10-day obstructed and 10-day denervated bladder ( ×3.5 for both, present study), and in diabetic bladders [10]. Neurotrimin is known to stimulate axonal growth, guidance of nerves to their targets, and be involved in synaptogenesis.
In recent years many studies have been published attempting to restore nerve function in urinary bladders with various lesions to the innervation. There has been some but limited success. In the following, we will review and reassess methods and approaches in the literature to restore bladder innervation after outlet obstruction and denervation. We intend to identify new directions for research and targets for therapeutic interventions.
MATERIALS AND METHODS
The present review is mainly a literature survey but contains also previously unpublished results from our bank of electron microscopy plates of obstructed and denervated female rat bladders, and bladder wholemounts stained for detection of acetylcholinesterase. We also have used unpublished microarray data from control, obstructed, and denervated rat bladders. These arrays have been used to characterize what happens at the molecular level during the growth of the obstructed bladder or denervated bladder and to check, in a systematic manner, the effect of obstruction and denervation for the mRNA expression of around 15,000 proteins. The microarrays have been used in other publications [3, 12, 13] and are publicly available (GEO accession numbers GSE47080 and 104540).
PREVIOUS RESULTS, NEW RESULTS, AND DISCUSSION
The symptoms of bladder innervation injury are secondary to either an increased spontaneous activity of the detrusor muscle cells causing an increased stiffness or overactivity of the bladder, or the contrary, an underactive bladder (UAB) with a large residual urine, and a decreased bladder sensitivity. It has been well demonstrated in different animal species that the obstructed bladder will show an increase in sensitivity to exogenously applied agonists and a reduction in sensitivity to intramural nerve stimulation (postjunctional supersensitivity). Morphological studies have shown an inverse correlation between nerve density and the duration of obstruction [14], leading to a dramatically decreased density of synapses in the long-term obstructed detrusor [15]. In animals [16, 17], but also in man [18, 19] the nerve damage is inhomogeneous. Urinary outlet obstruction can elicit a partial (patchy) denervation of the bladder, probably caused by e.g., ischemia in the bladder wall and stretch of the intramural nerves caused by the development of residual urine [16, 17, 19, 20]. Fusco et al. [21] summarized available evidence about BOO-induced morphological and molecular alterations occurring in the various compartments of the human bladder and concluded that the remodeling process involved multiple cellular compartments including not only detrusor muscle and nerves but also the urothelium, lamina propria, and detrusor extracellular medium, thus confirming what has been found in numerous animal studies.
In a series of acetylcholinesterase-stained wholemounts of the rat bladder the major nerve pattern of the obstructed bladders was found to be the same as in the control bladders (Fig. 1). The nerves were thicker, but the anatomical pattern of intramural nerves was the same as in the controls. No new nerves were observed in any of the obstructed bladders, and there were no intramural ganglia.
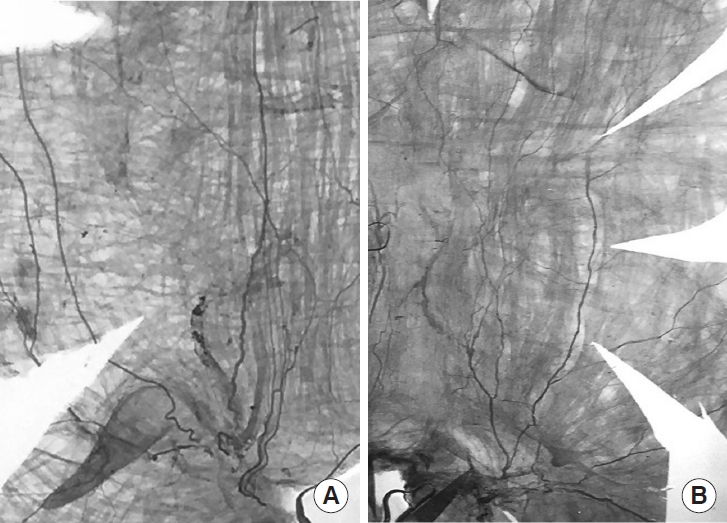
Wholemounts of acetylcholinesterase-stained ventral segment of a control bladder (A), and a 6-week obstructed bladder (B). Urethral orifice at the midbottom. The weight of the control and obstructed bladder was 70 mg, and 700 mg, respectively. The picture with the obstructed bladder is diminished 90% to give the same size as that of the control. No new nerves have developed in the obstructed bladder. Instead, there seems to be fewer nerves. No intramural ganglion cells can be seen. When the different magnifications are considered, it is obvious that the muscle bundles in the obstructed bladder are much larger than in the control. The amount of muscle in relation to the visible nerves is considerably higher in the obstructed bladder. The base of the left and right picture corresponds to 8.2 and 15.8 mm, respectively. The equatorial circumference was 36 mm (control), and 98 mm (obstructed). Both pictures are from the left side of the bladder. The photo of the control bladder is reversed to simplify the comparison.
Compensatory Mechanisms
Restoration of nerve function after injury can occur spontaneously or as the result of different types of intervention. The bladder innervation has a high degree of plasticity. This means that after a nerve lesion, surviving axons can sprout and take over the innervation from those axons that have degenerated [22]. This indicates that the capacity for restitution is high. There is also a high safety margin. Every muscle cell has at least one motor synapse, and there is connexion-dependent coupling between the cells making it possible for one active muscle cell to activate its neighbors. The bladder can lose up to 50% of its motor and sensory innervations without the development of residual urine [23].
Relief of Obstruction
The most obvious way of restoring nerve function of the obstructed bladder is of course to relieve the obstruction. After obstruction relief, there was a return toward control values of the physiological responses to exogenously applied agonists (acetylcholine and potassium) and electrical field stimulation [24]. There was also an increase in neuronal innervation which was more marked in the animals with a shorter period of obstruction. However, this relief of obstruction does not normalize the structural changes. Deobstructed bladders have characteristic gene expressions, structural morphologies, and functional properties that make them distinctly different from both control and obstructed bladders [3].
Ganglion Cell Differentiation
If the rat pelvic ganglia are removed or destroyed, the postganglionic parasympathetic axons in the bladder wall disappear completely causing a postganglionic denervation of the bladder. If the preganglionic nerves are cut, a decentralization of the bladder will occur but the ganglion cells and the postganglionic axons are left intact. The sensory axons from the bladder pass through the pelvic ganglia on their way to the spinal cord. Pelvic ganglionectomy thus also elicits a sensory denervation.
The nerves in the bladder wall contain axons and their supporting Schwann cells, surrounded by a fibrous sheath with collagen and fibroblasts. After unilateral ganglionectomy, the axons of the ipsilateral bladder nerves degenerate and disappear. The fibrous sheath and the Schwann cells do not degenerate, so the pattern of the distribution of the axon-free nerves is the same as in the normal bladder. If axons sprout, they can be led by the nerves to more distally located detrusor muscle cells. Fig. 2 shows a control ventral nerve and a corresponding nerve 3 months after ipsilateral ganglionectomy.
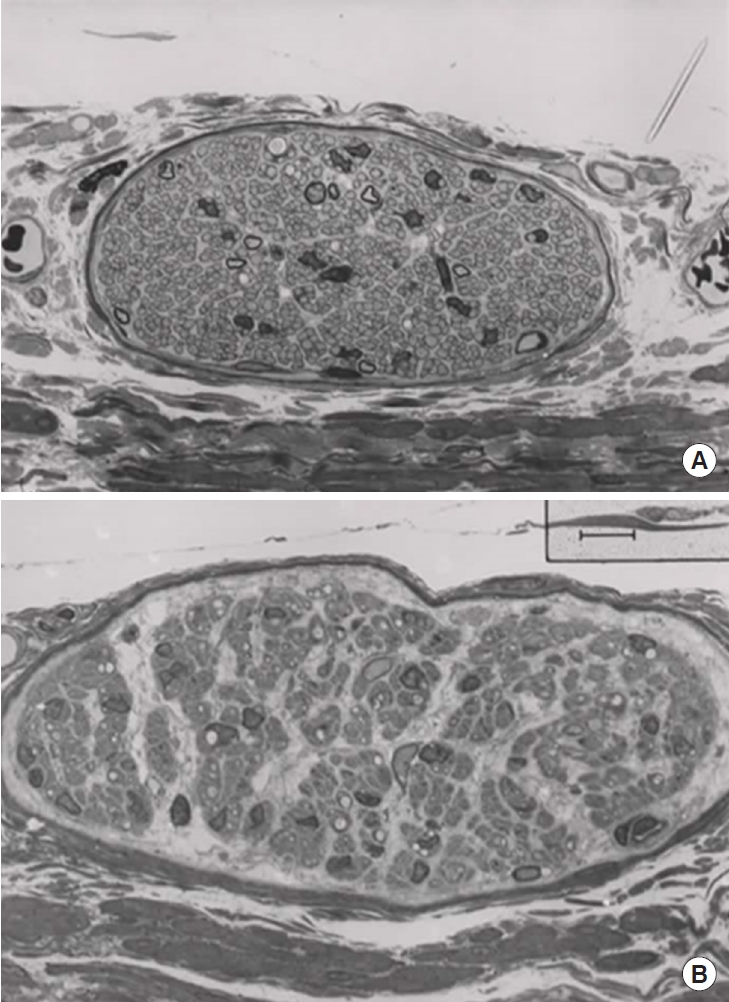
A ventral longitudinal nerve from a control bladder (A) and from a bladder 3 months after ipsilateral excision of the pelvic ganglion (B). The control nerve contains many Schwann cells, and unmyelinated axons to the detrusor muscle cells. A limited number of myelinated axons are sensory. In the lower panel the nerve still contains many Schwann cells after 3 months, but there is no sign of axonal sprouting. The surviving “axon-free” nerves probably have an important role in guiding the sprouting axons to their target, the smooth muscle cells. Calibration bar is 5 μm.
New ganglion cells can differentiate (Fig. 3) in the bladder wall from precursor cells along the intramural nerves [25]. A female rat pelvic ganglion contains about 2,600 ganglion cells [26]. If one pelvic ganglion is removed [25] about 50 new intramural ganglion cells seem to develop. This corresponds to about 1.9% of the number in the remaining ganglion. Whether the number of ganglion cells also increases in the remaining ganglion is not known. It is, however, known that the number of ganglion cells has increased to 3,995 [26] in adult female Brattleboro rats with hereditary diabetes insipidus. This suggests that the increased functional demand of the detrusor has led to the development of an increased number of ganglion cells. It can, however, not be ruled out that there is an age-dependent decrease in the number of pelvic ganglion neurons, and that this decrease is slowed down by the increased functional demand [27].
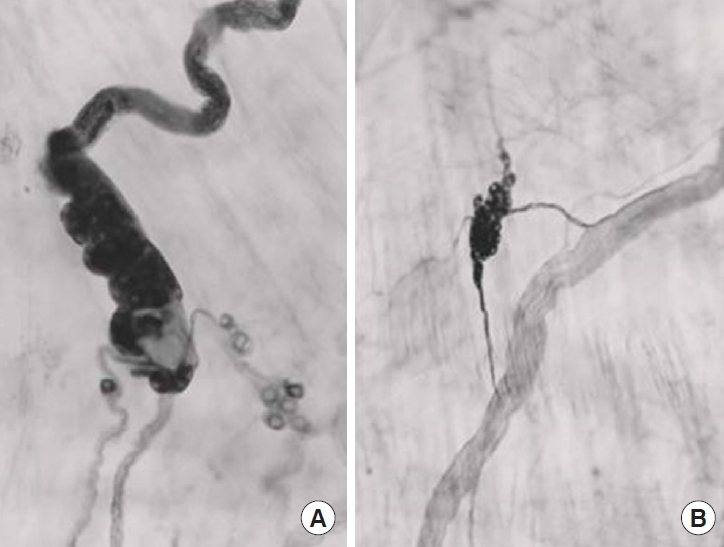
Ipsilaterally ganglionectomized bladder wall. New ganglion cells. Acetylcholinesterase staining. Panel A shows 2 axonfree (“denervated”) nerves coming from the bladder base and running distally (upwards). A newly developed cluster of ganglion cells is situated where these “empty” nerves coalesce, and the single nerve running distally (upwards) is filled with acetylcholinesterase staining axons. (B) An axon-free nerve is running in a distal direction (upwards). Axons from a nearby cluster of ganglion cells send 2 axon bundles to the nerve. The upper bundle then runs distally along the nerve. The size of the ganglion cells is about 25 μm.
Neurotrophin Stimulation
The axonal sprouting from cultured mouse pelvic ganglia in vitro is stimulated by the neurotrophins BDNF and neurotrophin-3 (NT3) whereas nerve growth factor (NGF) has little effect [11]. We have previously reported an increased level of BDNF mRNA in the obstructed bladder wall [3]. This increases BDNF level which then stimulates the axonal sprouting of parasympathetic intramural axons, and perhaps also the development of new ganglion cells. Preliminary experiments (Uvelius and Gabella) suggest that superior cervical ganglion (SCG) neurons (transmitter noradrenaline) do not sprout when transplanted into the bladder wall. The SCG neurons have TrkA receptors, instead of TrkB and C as the pelvic ganglion cells and would therefore need NGF, which is low in the bladder, as its neurotrophin. Neurotrimin, although not a neurotrophin [10] has an increased mRNA in obstructed bladders. If it was possible to increase the concentration of BDNF, NT3, or neurotrimin in the wall of the obstructed bladder one would expect more sprouting axons from pelvic ganglion neurons.
Ganglion Cell Transplantation
An interesting approach to the restoration of damaged bladder denervation was described by Gabella and Uvelius [28] who transplanted a large portion of the pelvic ganglion of adult female rats into the wall of the urinary bladder of the same animals. Since the rat urinary bladder does not normally contain any intramural ganglion cells this makes it a good model to study bladder denervation and decentralization, giving experimental advantages over bladders that, like the human, have intramural ganglia. Gabella and Uvelius [28] found that the general architecture of all the transplanted ganglia was preserved, and that the vascularization was re-established. The nerves grew out of the transplant and connections with the original intramural nerves. All the synapses degenerated at the time of transplantation, but new synapses began to reappear on the ganglion neurons and most neurons survived for the full length of the experiments (4 months). The authors suggested that their results showed that homotransplants of pelvic neurons into the bladder have a high rate of success, in terms of survival, maintenance of fine structure, growth, and reconnections. In addition, these neurons of adult organisms display plastic and regenerative abilities. The bladder innervation has a high degree of plasticity. This means that surviving axons can sprout and take over the innervation from those nerves that have degenerated. Gabella and Uvelius [28] did not investigate the functional status of the bladders after transplantation. If bladder function can be restored, this approach may be possible to apply in humans: parasympathetic ganglion cells could be injected into the bladder wall through a cystoscope in the same manner as Botox injections. A future possibility would be to use ganglion cells with different transmitters.
Stem Cell Therapy
An attractive way to improve function in an obstructed and or denervated bladder would be stem cell therapy [29-33]. Studies of stem cells injected into the bladder have demonstrated positive effects in experimental voiding dysfunctions such as detrusor overactivity with or without bladder outflow obstruction (BOO). Most studies on the effects of stem cells on bladder dysfunction have been performed on the rat BOO model [31, 34, 35].
It is obvious that stem cell treatment can affect the remodeling of the bladder that occurs after the initiation of obstruction in the rat BOO model, and that this treatment may have a beneficial effect against both morphological and functional changes. In many of the animal studies performed, the authors speculated on the use of mesenchymal stem cells (MSCs) as a new treatment to prevent the morphological and functional effects of BOO in humans. However, if MSC treatment has positive effects also in human BOO, relevant questions are (1) When should treatment be instituted?, (2) can cell therapy reverse established morphological changes? In most of the BOO studies cited above, stem cells were given early in the remodeling process initiated by the urethral obstruction, i.e., at times points when the process is still dynamic. Studies on the clinical application of MSCs in humans with various diseases are available. However, the effects of stem cell therapy for bladder dysfunction, e.g., in patients with OAB, BOO-associated detrusor overactivity, or patients with UAB have so far not been published.
Anti-Inflammatory Therapy
The progressive urinary dysfunction associated with BOO has been known for over a decade to be the result of an ongoing inflammatory response triggered by at least 3 interrelated insults: (1) high pressure, (2) excessive stretch, and (3) hypoxia/reperfusion (oxidative stress), all of which occur repeatedly with each micturition cycle as the bladder contracts with greater force to overcome the outflow resistance [21]. Of particular interest in this context are the nucleotide-binding oligomerization domain-like receptors, especially the best-studied family member, NLR family pyrin domain containing 3 (NLRP3). Hughes et al. [36] studied the importance of NLRP3 in a chronic setting to determine whether the NLRP3 inhibitor glyburide, affected inflammation, voiding dysfunction, bladder decompensation, and nerve density in long-term obstruction. The nerves they measured were identical to those seen in Fig. 1 but with the bladders cut at a right angle to their long axis. Note that they did not measure the density of synapses in their light microscopy study, and a decreased nerve density does not necessarily indicate a decrease in the density of synapses, i.e., denervation.
Hughes et al. [36] found 7 nerves per 106 μm2 in their control rat bladders. Sadegh et al. [15] found, in similar bladders, 1 synapse per 100 μm2 (104 synapses per 106 μm2). They used electron microscopy which has a much higher resolution and makes it possible to directly count synapses. In the obstructed bladders Hughes et al. [36] found 2 nerves per 106 μm2, whereas Sadegh et al. [15] found 0.1 synapse per 100 μm2 (103 synapses per 106 μm2). This means that obstruction decreases nerve density by a factor 3.5, and synapse density by a factor of 10. As Hughes et al. [36] defined nerves this means that there are about 1,400 synapses per nerve in control bladders and 500 in the obstructed. The measurements of nerve densities are thus not a direct function of the synapse densities but relative comparisons between nerve densities seem possible.
Hughes et al. [36] noted functional improvement after glyburide treatment, e.g., the decrease in voiding efficiency in the chronic BOO rats was prevented. Of particular interest was that the reduction in nerve density apparent with BOO was attenuated with glyburide. The authors concluded that their results suggested a critical role for NLRP3 in mediating bladder decompensation and nerve density during chronic BOO. It is not known, however, if glyburide has any effect on the density of synapses. We found in our arrays that NLRP3 mRNA increased by 90 percent in 10-day denervated bladders but was unaffected by obstruction.
It is unclear whether Cox-2 is involved in the changed nerve density following obstruction. Cox-2 mRNA is, however, increased almost 5 times in 10-day obstructed bladders but was unaffected by 10-day denervation. This fits well with the results in the study by Park et al. [37]. They studied the effect of acute total obstruction in the mouse bladder and found that 6 hours of obstruction induced a pronounced increase of Cox-2 mRNA caused by the stretch of the detrusor smooth muscle cells. NS-398, a Cox-2 inhibitor could prevent this increase.
We speculate that NLRP3 is increased in the denervated bladders by neurogenic inflammation caused by the degenerating intramural axons, whereas the increased Cox-2 mRNA is caused by the stretch of the bladder wall in our partially obstructed bladders.
SUMMARY
Relief of obstruction, e.g., transurethral resection of the prostate (TURP) usually leads to relief of symptoms, and the patient feels cured. If symptoms of underactivity persist after the normalization of outflow conditions, the treatment options are limited, and include both re-TURP and clean intermittent catheterization. In the present review, we speculate what will become the future options.
There will be both local and systemic treatment. As of today, the standard local treatment will aim at reducing outlet obstruction. Other local treatments might be transplanting stem cells or mature bladder ganglion cells into the bladder wall. The literature shows that there has been some progress. It is possible to transplant ganglion cells experimentally into the bladder. The ganglion cells survive and sprout into the surrounding tissue. So far, it is not known if there are functional connections (synapses) between the axons of the transplanted ganglion cells, and the detrusor smooth muscle cells. Stem cell transplantation has been performed successfully but with a result opposite to that of ganglion cell transplantation. Transplantation of stem cells improves the function of the detrusor, but it has been difficult to demonstrate the transplanted stem cells, or the cells derived from them.
Neurotrophins or neurotrimin might be injected in the bladder wall to increase the sprouting of existing ganglion cells, or cells that have been injected into the bladder wall.
Bladder outlet obstruction, persisting underactivity, and decreased nerve density are often combined with inflammatory activity of the lower urinary tract. NLRP3 and its mRNA as well as Cox-2 mRNA are increased in obstructed bladders. Systemic treatment with the NLRP3 inhibitor glyburide normalized nerve density, and, to some extent, bladder function. It is unclear whether Cox-2 is involved in the changed nerve density following obstruction, but Cox-2 mRNA increases almost 5-fold in the obstructed bladder.
CONCLUSIONS
Without being too speculative future therapy against bladder underactivity remaining following relief of outlet obstruction could be either a systemic therapy, perhaps by anti-inflammatory drug treatment, or a local using neurotrophins, ganglionic or stem cells, perhaps by injection into the bladder wall as Botulinum toxin is injected today.
Notes
Grant/Fund Support
This study received no specific grant from any funding agency in the public, commercial, or not-for-profit sectors.
Research Ethics
The study does not contain any new animal experiments. Ethical permits for the original experiments were applied for at the Swedish Board of Agriculture and approved by the Malmo/Lund Ethical Committee for animal experiments (approval numbers M 4–16, M 104–15, and M 40–13).
Conflict of Interest
No potential conflict of interest relevant to this article was reported.
AUTHOR CONTRIBUTION STATEMENT
· Conceptualization: BU, KEA
· Data curation: BU, KEA
· Formal analysis: BU, KEA
· Funding acquisition: BU, KEA
· Methodology: BU, KEA
· Project administration: BU, KEA
· Visualization: BU, KEA
· Writing: BU, KEA
Acknowledgements
We acknowledge the long-lasting cooperation with Professor Giorgio Gabella.