• HIGHLIGHTS
- A rat model of bladder outlet obstruction (BOO) demonstrated reduced angiopoietin expression.
- Visceral ischemia and fibrosis are associated with BOO progression.
INTRODUCTION
In aging males with benign prostatic hyperplasia (BPH), bladder outlet obstruction (BOO) frequently occurs [1,2]. Over time, BOO can lead to bladder decompensation and impaired visceral function [3]. Understanding the timeline of disease processes impacting the bladder in this scenario could aid in determining the optimal window for surgical repair.
Animal models of BOO are frequently used to explore the disease mechanisms associated with bladder problems in individuals suffering from BPH. In in vivo experiments, BOO can induce early hypertrophy of the detrusor muscle and increase intravesical pressure to maintain steady urine flow [1-3]. This process can compromise the blood supply to the detrusor muscle, leading to ischemia, which plays a significant role in abnormal detrusor function. As a consequence, metabolism may shift towards anaerobic mechanisms [4,5].
The transcription factor hypoxia-inducible factor 1-alpha (HIF1-α) is a crucial mediator in mammalian cells, becoming active only under conditions of cellular hypoxia [6,7]. As an oxygen-dependent transcriptional activator, it has the ability to activate genes involved in glucose transport and metabolism. It is also responsible for amplifying the expression of genes known to contribute to cell survival and programmed cell death [8]. In particular, the upregulation of HIF1-α can stimulate the expression of proangiogenic gene products, including vascular endothelial growth factor (VEGF) [9].
BOO can lead to compensatory changes within the bladder. These changes may provide a deeper understanding of the disease processes underlying the condition, thereby assisting in determining the optimal timing for bladder repair in patients with BOO. The expression of HIF1-α, VEGF/VEGF receptor, and angiopoietin/angiopoietin receptor might serve as potential markers of tissue hypoxia and visceral adaptation. The aim of the current study was to measure the mRNA levels of these factors in a rat BOO model, in order to investigate the bladder’s initial response to injury caused by BOO.
MATERIALS AND METHODS
Rat BOO Model
All protocols in this study adhered to the National Institute of Health Guide for the Care and Use of Laboratory Animals. The study received approval from the Institutional Animal Care and Use Committee of the clinical facility. The BOO model used in this study was developed by replicating the study protocol previously conducted by the authors [10,11].
Twenty female Sprague-Dawley rats, aged 6 weeks, were divided into 4 groups, with each group containing 5 rats. Group 1 (n=5) served as the control, while group 2 (n=5) underwent sham surgery. Groups 3 (n=5) and 4 (n=5) underwent BOO surgery and were monitored for 1 week and 8 weeks, respectively. BOO surgery was performed on the relevant rats under isoflurane (BK Pharm, Goyang, Korea) anesthesia. A skin incision was made in the lower abdomen, followed by urethral dissection. Subsequently, 4-0 silk sutures were used to encircle the urethra, with a 1-mm external diameter metal rod placed outside the lumen. Once the sutures were placed, the rod was removed and the abdominal wound was closed. For infection prophylaxis, cephalosporin (Flomoxef, BK Pharm, Goyang, Korea, 10 mg/kg per day) was administered interperitoneally.
Measurement of Voiding Function
Eight weeks after BOO surgery, the voiding response was evaluated in all rats by inserting a PE-50 catheter into the bladder. After the administration of an isoflurane anesthetic, a midline abdominal incision was made to expose the bladder. A collar was formed by warming the bladder end of the catheter, which was then positioned in the bladder via a tiny incision in the bladder dome and held in place by a suture. The opposing catheter end was threaded through the subcutaneous tissue to exit the skin. The wound was closed by suturing muscular and dermal layers. Anesthesia was then terminated.
The rats were housed in a restraining cage for 6 hours, which included a 2-hour recovery period following the surgical anesthesia. The dimensions of the cage were designed to allow the rats to crouch comfortably, but not to turn around. Once the rats had fully recovered from the isoflurane anesthesia, a pump was connected to the bladder catheter via a T-stopcock. This setup facilitated the continuous infusion of physiological saline, maintained at room temperature, into the bladder at a rate of 0.04 mL/min. The pump was also linked to a pressure transducer. The saline infusion triggered multiple voiding responses in the rats. The urination reflexes induced by volume loading were studied in the restrained rats. The parameters that were recorded included the intravesical pressure necessary for micturition, the pressure threshold (PT), the maximal voiding pressure (MVP), and the intercontraction interval (ICI). The ICI was defined as the time elapsed between voiding episodes or reflex bladder contractions.
Histology and Immunohistochemistry
After cystometry, the rats were euthanized via an intracardiac infusion of 100 mL of cold saline and 100 mL of 4% paraformaldehyde in phosphate-buffered saline, while under anesthesia. The bladder was subsequently extracted from each rodent and preserved in 4% paraformaldehyde for 48 hours. The viscera were then cryoprotected using 30% sucrose for 72 hours. Subsequently, a cryostat (Leica CM 1900, Leica, Wetzlar, Germany) was employed to cut sections of 10-μm thickness. These sections were then subjected to fluorescent staining and immunofluorescence processing. The samples were examined using an Olympus laser confocal fluorescence microscope.
The samples were subjected to immunostaining to detect the presence of von Willebrand factor (vWF) (1:200, Abcam, Cambridge, UK). Antigens were extracted following a 10-minute warming period with 9mM sodium citrate at a pH of 5. To inhibit intrinsic peroxidase activity, a peroxidase-blocking reagent (Dako, Carpinteria, CA, USA) was utilized. Free-floating tissue sections were incubated overnight at 4°C using a mixed primary antibody solution for vWF. Subsequently, Alexa Fluor 488-conjugated anti-mouse IgG (1:200, Molecular Probe, Eugene, OR, USA) and Alexa Fluor 594-conjugated anti-rabbit IgG (1:200, Molecular Probe) were added, followed by an additional 60-minute incubation at room temperature. A negative control specimen was prepared in the same manner, but without the inclusion of primary antibodies. The stained samples were examined using an Olympus laser confocal fluorescence microscope. Quantitative analysis of the histological examinations was conducted using an image analyzer system (National Institutes of Health Image J 1.34, http://rsbweb.nih.gov/ij/).
mRNA Expression
As stated above, the rodents were sacrificed after cystometry was completed. The bodies of the bladders were removed and dissected away from any attached soft tissues prior to weighing. They were placed directly into a freezer in liquid nitrogen until additional biochemical and molecular biological assessments were required.
The Trizol reagent (Invitrogen, Carlsbad, CA, USA) was used to extract total DNAs from cells and samples. Real-time polymerase chain reactions (PCRs) were carried out. The threshold cycle value was the basis for assaying relative gene expression. Normalization was performed against β-actin as a housekeeping gene. Data averaged over 3 experiments are presented. The PCR procedure comprised the following stages: 95°C for 10 seconds, and 40 cycles of 95°C for 5 seconds and 60°C for 30 seconds. Lastly, melting curve analysis over 60°C–95°C was carried out.
Real-Time PCR Array for Hypoxia-Related Genes
To isolate and extract total RNA, an RNA extraction kit (RNeasy Mini Kit, QIAGEN, Hilden, Germany) was used following the manufacturer’s instructions. The extracted RNA solution was treated with RNase-free DNase (QIAGEN, Hilden, Germany) to ensure there was no DNA contamination. Using an RT2 First Strand Kit (QIAGEN) and 2 μg of total RNA, the initial cDNA strand was synthesized in accordance with the manufacturer’s recommendations. This was then combined with RT2 quantitative PCR master mixes. Small quantities of this mixture were placed in a 96-Well RT2 Profiler PCR Array for rat hypoxia (QIAGEN). This array included SYBR green-optimized primer assays for a carefully selected panel of genes relevant to the pathway, consisting of 84 genes associated with the hypoxic response and 12 genes serving as a reference for the quality of loading and cDNA. The real-time PCR assay was conducted by heating the plate for 10 minutes at 95°C, followed by 40 cycles of 15 seconds at 95°C and 60 seconds at 60°C. Quality assurance was provided by genomic DNA, reverse transcription, and positive PCR controls. β-actin served as the housekeeping gene. Any changes in gene expression were represented as a factor of increase or decrease, with a cutoff value of changes over a factor of 2 defining upregulated or downregulated expression. The primers used for real-time PCR for hypoxia are listed in Table 1.
RESULTS
Body and Bladder Weight
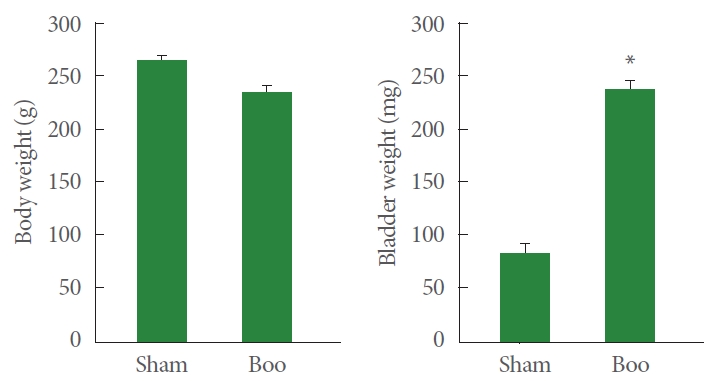
The mean body mass of the rats in the sham surgical group and groups receiving BOO were equivalent (266±3.3 g and 235±7.1 g, respectively). The rats that underwent sham surgery had a significantly lower bladder weight (86.9±8.5 mg vs. 242.2±7.8 mg) than those that underwent a BOO procedure (242.2±7.9 mg) (P<0.05) (Fig. 1).
Impaired Bladder Contractility
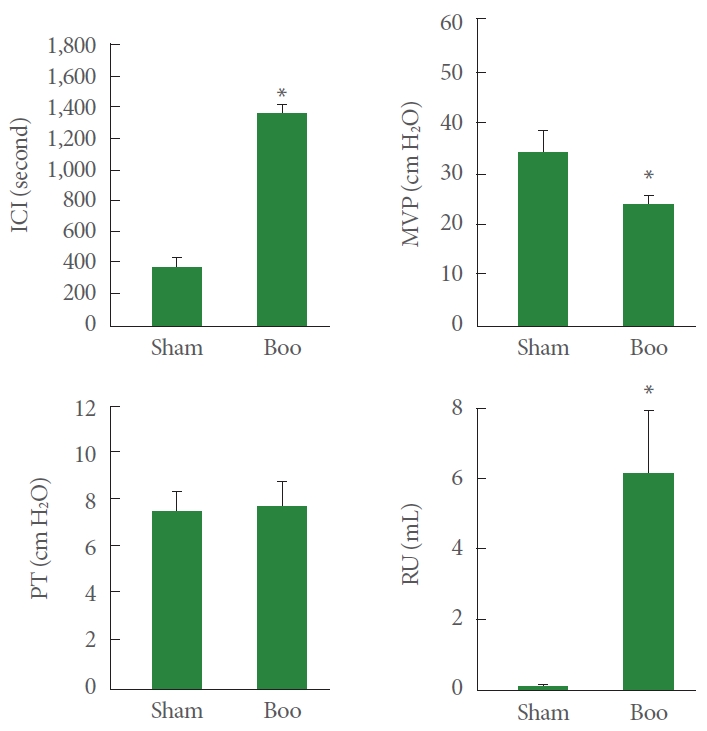
In rats that underwent BOO, the ICI was considerably higher (1,384±214 seconds) than in the sham surgical cohort (390±58 seconds, P<0.05). The PT values for the 2 groups were comparable (7.8±0.7 cm H2O in the sham surgical group and 8.0±0.8 cm H2O in the BOO cohort). The BOO group showed a lower MVP than the sham surgery group (23.7±4.1 cm H2O vs. 34.7±7.4 cm H2O) with elevated RU (6.4±1.5 mL vs. 0.1±0.08 mL) (Fig. 2).
mRNA Expression in the Bladder of the BOO Group
Relative to the control group, HIF-1α mRNA expression levels were similar in rats that underwent BOO at week 1, but increased at 8 weeks following the BOO procedure (P<0.05). VEGFA and VEGFR2 mRNA expression levels at 1 week and 8 weeks following BOO were similar to those in the control group.
Compared with rats in the sham surgery group, the group at 1 week after BOO demonstrated higher levels of angiopoietin-1 (Ang-1), Ang-2, Ang-4, and Tie2 mRNA expression (Fig. 3). However, these levels were diminished at 8 weeks after BOO (P<0.05).
Histopathology
Fewer vWF-positive blood vessel configurations were identified on immunofluorescence staining in rats that underwent BOO (P<0.05) (Fig. 4).
Hypoxia Signaling Pathway Analysis by PCR Array
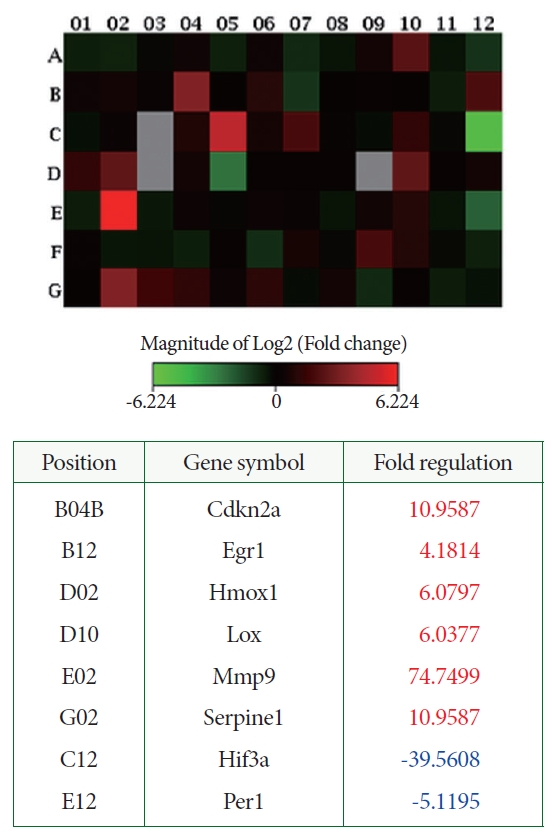
Hif3a (-39.6) and Per1 (-5.1) were among the HIF and cotranscription factors. Additional HIF interactors included cyclin-dependent kinase inhibitor 2A (Cdkn2a, 11.0). Early growth response 1 (Erg1, 4.2), matrix metalloproteinase 9 (Mmp9, 74.7), serpin peptidase inhibitor, clade E, member 1 (Serpine1, 11.0), lysyl oxidase (LOX, 6.0), and heme oxygenase 1 (Hmox1, 6.1) were related to angiogenesis (Fig. 5).
DISCUSSION
BOO can lead to mural thickening of the bladder and hypertrophy, which can improve bladder stability and facilitates adaptation to the requirement to void against an increase in resistance in the urethra caused by the obstruction [12,13]. Mural thickening creates a cycle of ischemia and reperfusion during the urination cycle [14,15]. Hypoxia in the bladder has been observed in the smooth detrusor muscle and bladder subserosa [16]. This oxygen deficit is associated with impaired contractile function and metabolic abnormalities in the smooth muscle.
In this study, BOO significantly affected the mass of the bladder (Fig. 1). BOO compromises mural perfusion in the bladder. Bladder contractions can result in reduced oxygen tension, which is then reversed after the bladder is emptied [14,17]. The viscera can adapt if BOO persists for only a short period. However, in the context of ongoing BOO, repeated episodes of hypoxia can lead to progressive functional impairment and bladder decompensation.
It has been established that a decrease in detrusor perfusion, observed immediately following BOO, correlates with the degree of bladder decompensation [17]. This decompensation occurs as the detrusor muscle undergoes repetitive phases of ischemia and reperfusion, leading to the production of reactive free radicals. Furthermore, the activation of specific phospholipase and protease enzymes can contribute to the membrane damage observed at both cellular and subcellular levels [18].
In the present study, we observed increases in ICI and RU, and a decrease in MVP (Fig. 2). Rapid alterations in the bladder’s configuration and functionality are initiated following the onset of BOO. Initially, the MVP increases to maintain a steady urine flow, while the ICI decreases [1-3]. Significant bladder decompensation was induced following BOO, resulting in the parameter changes as described. In a system where BOO exists, the process of filling and emptying the bladder can trigger a cycle of ischemia and reperfusion within the viscera. This cycle can rapidly alter the oxygenation status of a tissue [19]. For hypoxia to be rectified, angiogenesis is essential. The term angiogenesis refers to the formation of new arterial and venous vessels. These may originate from pre-existing vasculature (for instance, through the creation of de novo vessels from current vascular conduits or by the division of these conduits), or from the formation of neovascular vessels as a result of sprouting angiogenesis [19,20].
Earlier studies have documented a close link between low oxygen levels and HIF-1α [21-23]. In mammalian cells, the transcription factor HIF-1α is responsible for the hypoxia response. This mediator is only functional under conditions of cellular hypoxia [6,7]. As an oxygen-dependent transcriptional activator, HIF-1α can enhance the activity of genes associated with glucose transport and metabolism. It can also trigger the upregulation of a gene that aids in maintaining cellular viability [8]. The HIF-1α transcription factor is the primary initiator of the hypoxia signaling pathway when oxygen availability is reduced. A variety of biological activities, such as angiogenesis, hematopoiesis, and the maintenance of vascular tone to ensure the delivery of blood and oxygen to tissues, are regulated by target genes that respond to low oxygen levels. In the rat model used in this study, BOO resulted in a minor change in HIF-1α levels after 1 week. However, after 8 weeks, a significant increase in HIF-1α levels was observed. This suggests that short-term BOO (e.g., 1 week) does not lead to bladder hypoxia, while long-term BOO (e.g., 8 weeks) does have hypoxic effects (Fig. 3).
It has been observed that alongside HIF-1α in BOO, VEGF levels also increase [24,25]. VEGF, which is an angiogenic factor and a diffusible mitogen specific to endothelial cells, increases the permeability of the vasculature, leading to substantial neovascular formation.
VEGFA is a prominent member of the VEGF family. Endothelial cells (ECs) are the only cells that express VEGFR2. This factor is potentially critical for the differentiation of ECs and the formation of blood vessels [24,25]. The modest alteration in HIF-1α levels observed in this study at week 1 failed to cause an increase in VEGF. Interestingly, no elevation in VEGF levels was seen at week 8 despite increased HIF-1α levels (Fig. 3). VEGFR2 expression remained relatively constant in rats that underwent BOO, both at 1 week and 8 weeks after BOO. The duration of BOO was sufficient to imply that these results indicated a failure of VEGF and VEGFR to respond to the increase in HIF-1α.
Angiogenesis is controlled at specific points by Ang-1 and VEGF. The latter promotes the sprouting and development of vascular conduits, whereas the former contributes to the maturation and remodeling of blood vessels [26]. VEGF influences the formation of new blood vessels, but the conduits it creates are structurally immature. This is evident in their lack of uniform appearance, tortuosity, dead ends, and disordered arrangement of venules, arterioles, and capillaries. Functionally, these conduits are also deficient, as they leak blood cells, which can lead to increased interstitial pressure.
ECs are responsible for establishing the vascular system, working in conjunction with auxiliary mural cells such as small capillary pericytes and smooth muscle cells from larger caliber vessels. These cell types work in close collaboration during the processes of neovasculature sprouting, cell migration and replication, and blood vessel maturation. They aid in the formation of new vessels that interconnect to form a well-branched network of conduits. This network facilitates the transport of fluids, nutrients, gases, hormones, and cells within the circulatory system throughout the body of a vertebrate organism. The remodeling and maturation of vessels, as well as the recruitment of mural cells during vasculature homeostasis in adults, have been shown to rely on the tyrosine kinase with immunoglobulin and EGF homology domains (Tie), angiopoietin receptor system [27]. In pericytes and smooth muscle cells, low oxygen levels and VEGF have been found to increase the expression of Ang-1. This, in conjunction with VEGF expression, can facilitate the development of well-differentiated vasculature. In mice expressing VEGFA alone, the Ang/Tie system was observed to counteract vessel leakage and permeability. The signaling function of the Ang/Tie system appears to act as a supplementary mechanism in the presence of VEGF and VEGFR.
Ang-2 is considered to have only partial agonistic activity due to its lower affinity for Tie2 compared to Ang-1. The role of Ang-3 and Ang-4 in neovascular formation related to disease has not yet been thoroughly investigated [28].
The current data demonstrated that, compared to rats that underwent sham surgery, rats 1-week post-BOO exhibited elevated levels of Ang-1/Tie and VEGF, as well as Ang-1, Ang-2, and Ang-4. The expression levels of these factors decreased in rats after 8 weeks of BOO (P<0.05) (Fig. 3). Tie2 followed a similar pattern (P<0.05) (Fig. 3). Survival under the low bladder oxygen levels induced by BOO necessitates the stimulation of angiogenesis to increase VEGF/VEGFR and Ang/Tie levels. There was no immediate increase in the former. However, HIF-1α expression was enhanced, and a decrease in angiopoietins along with their receptors was observed 8 weeks after BOO.
When the bladder is chronically underactive, it exhibits decompensation. Hypoxia can trigger an inappropriate reaction. Rather than VEGF/VEGFR promoting angiogenesis, Ang/Tie responds to the low oxygen tension to ensure survival. These findings suggest that bladder angiogenesis may be promoted to compensate for the new metabolic condition that arises 1 week after BOO. However, this response was diminished at 8 weeks after BOO. Hypoxia has been observed in the smooth detrusor muscle and the bladder subserosa [16]. The BOO cohort exhibited fewer vascular conduits and a lower vessel density compared to rats that underwent sham surgery or controls, as evidenced by vWF immunofluorescence staining (Fig. 4). Low oxygen tension resulting from the surgical procedure was noted.
The 96-Well RT2 Profiler PCR Array for rat hypoxia (QIAGEN) includes SYBR green-optimized primer assays for a carefully selected panel of genes pertinent to the pathway. This array covers 84 genes linked to the hypoxic response. In our recent study, we analyzed bladder samples taken from rats 8 weeks after BOO. We found that the expression of HIF3a, a gene known to negatively mediate gene expression under low oxygen conditions, was downregulated by a factor of 39.6 (Fig. 5). This suggests that Hif3a may have a positive influence on gene expression in hypoxic conditions. In the same group of rats, we observed a downregulation of Per1, which is associated with HIF and cotranscription factors, by a factor of 5.1 in bladder samples. We also noted an upregulation of Cdkn2a, a secondary HIF1 interactor, by a factor of 11.0. Furthermore, the genes erg1, Serpine 1, and Hmox1 were upregulated by factors of 4.2, 11.0, and 6.1, respectively. This specific gene expression pattern could potentially explain the molecular disease processes observed in a decompensated bladder with BOO, which is associated with the activation of the HIF-1 pathway.
Many disease mechanisms are considered to result from low oxygen tension, such as the deposition of extracellular matrices and changes in smooth muscle function [29]. LOX, an extracellular enzyme, can facilitate covalent crosslinking between type I and type III collagen fibers, thereby enhancing the stiffness and stability of extracellular collagen [30]. Upregulation of LOX by a factor of 6.0 was observed in the bladders of rats at 8 weeks after BOO. In angiogenesis and wound reparation, degradation of the extracellular membrane is caused by matrix metalloproteinases (MMPs) [31]. Upregulation of Mmp9 by a factor of 74.7 was seen in the current work, suggesting a positive influence of Mmp9 on angiogenesis within the bladder at 8 weeks after BOO. These results established that sprouting of the endothelium, enhanced permeability of the vasculature, tissue MMP expression, and ultimately, matrix digestion, are necessary for cellular migration. This research provides evidence regarding changes in the pathways of hypoxia-related genes in response to BOO.