Decreased Expression of Urethral Caveolin-1, −2, and −3 in the Rat Model of Overactive Bladder: Potential Mediator of Functional Interaction of Urethra and Urinary Bladder
Article information

Abstract
Purpose
To investigate the effect of detrusor overactivity (DO) on the urethral expression of caveolin (CAV)-1, −2, and −3 of urethra in an animal model of cyclophosphamide (CYP)-induced cystitis rat.
Methods
Female Sprague-Dawley rats were divided into the control group (n=20) and the cystitis group (n=20). Cystitis was induced by intraperitoneal injection of CYP (200 mg/kg). An urodynamic study was done 3 days after the CYP injection to measure functional change of the urinary bladder and urethra. Cellular localization and expression of CAV-1, −2, and −3 in the rat urethra were determined by immunohistochemistry (IHC) and Western blot.
Results
Urodynamic experiments demonstrated a decreased contraction interval in the cystitis group compared to the control (3.9±1.0 minutes vs. 6.6±1.2 minutes, P<0.05). Conversely, contraction pressure increased significantly in the cystitis group compared to the control (22.4±0.7 mmHg vs. 11.5±0.4 mmHg, P<0.05). The urethral pressure was decreased in the cystitis group compared to the control (4.05±2.5 mmHg vs. 5.8±2.8 mmHg, P<0.05). The IHC and Western blot data showed that CAV-1, −2, and −3 expression decreased significantly in the cystitis group compared control group (P<0.05).
Conclusions
The decreased urethral CAV-1, −2, and −3 in the DO rats suggests that CAVs might be related with the functional change of urethra in association with DO of urinay bladder.
• HIGHLIGHTS
- The current study was designed to investigate the functional change and the expression of caveolin (CAV)-1, −2, and −3 in the urethra and to suggest the potential implications of CAVs on urethral function in a detrusor overactivity (DO) animal model of cyclophosphamide (CYP)-induced cystitis rat. No study, to date, has yet evaluated the expression of CAVs in the urethra when the pathological changes of the urinary bladder occur.
- Our study indicates that the functional change of urethra and the decreased expression of urethral CAV-1, −2, and −3 was observed in the animal model of DO by CYP-induced cystitis rat.
- These data suggest that urethral CAVs may, in part, be involved with the functional change of urethra in association with DO of urinary bladder.
INTRODUCTION
Caveolins (CAVs) are flask-shaped small invaginations of the plasma membrane that are thought to play an important role in cell surface signaling, endocytosis, and intracellular cholesterol transport [1]. These structures are present in most cells but are abundant in endothelial cells, adipocytes, type 1 pneumocytes, fibroblasts, and smooth and striated muscle cells. The CAV family includes 3 members—CAV-1, −2, and −3—that have different expression patterns specific to a particular cell-type [2]. CAV-1 is predominantly expressed in adipocytes, vascular endothelial cells, fibroblasts, smooth muscle cells, and a variety of epithelial cells [3]. CAV-2 is tightly co-expressed with CAV-1, whereas CAV-3 is highly expressed in muscle cells [4,5]. CAVs may be involved in the pathogenesis of various diseases by dysregulating signal transduction pathways [6,7].
It has been reported that the CAVs is associated with physiological function of the urinary bladder. We suggested previously that CAV-1 might be involved in urinary bladder signaling activity and play a role in bladder dysfunction in a cyclophosphamide (CYP)-induced rat cystitis model showing detrusor overactivity (DO) [8].
However, it seems like that the urethral function is tightly associated with the physiologic alteration of the urinary bladder. Wise et al. [9] investigated the urethral response in patients with overactive bladder and reported that urethral instability occurs in 42% of patients with these symptoms. Chaliha et al. [10] assessed the effects of DO on the functional change of the urethra using the urethral profilometry and suggested that urethral function is strongly affected by the presence of abnormal detrusor activity.
Until now, studies on the association between lower urinary tract and CAV have been largely confined to the bladder. However, a study was conducted on the relationship between urethralfunction and CAV using CAV knock out (KO) mice [11]. According to the study, cellular localization and protein expression of CAV were detected in the mouse and urethra. And CAV expression was reduced in CAV KO urethra and maximal urethral contraction was reduced in CAV KO mice. Sancho et al. [12] analyzed the changes in the expression and distribution of nitric oxide synthase (NOS), and the alterations in nerve-mediated contractility in the bladder and urethra of CYP-treated rats. They found transient downregulation of neuronal NOS expression and a progressive upregulation of endothelial NOS and qualitative changes in mRNA expression were also observed in the bladder and urethra.
The current study was designed to investigate the functional change and the expression of CAV-1, −2, and −3 in the urethra and to suggest the potential implications of CAVs on urethral function in a DO animal model of CYP-induced cystitis rat. No study, to date, has yet evaluated the expression of CAVs in the urethra when the pathological changes of the urinary bladder occur.
MATERIALS AND METHODS
Experimental Model
Female Sprague-Dawley rats (age: 14 weeks, weight: 250–300 g, n=40) were divided into control (n=20) and experimental cystitis group (n=20). The cystitis group was induced by intraperitoneal injection of CYP (200 mg/kg) (Sigma Chemical Co., St. Louis, MO, USA) [13]. The control group was intraperitoneally injected with saline. The rats were premedicated with xylazine (2.2 mg/kg, intramuscularly [IM]) (Sigma Chemical Co.) and anesthetized with a zolazepam/Tiletamine cocktail (4.4 mg/kg, IM) (Sigma Chemical Co.). Cellular localization and expression of CAV-1, −2, and −3 were determined by immunohistochemistry (IHC) and Western blot of the rat urethra. All rats were fed a standard diet.
Urodynamic Study
Urodynamic studies were conducted 3 days later to measure contraction interval and contraction pressure. The rats (n=5/group) were anesthetized with a subcutaneous injection of urethane (1.2 g/kg) 3 days after the CYP injection. To monitor the vesical and urethral pressure, the rats were prepared using the method of Bae et al [14]. The bladder was exposed via a suprapubic midline incision. The contraction pressure and contraction interval of the urinary bladder, and the urethral pressure were monitored using a custom designed triple-lumen catheter, which consisted of an outer catheter (8-french cut-down tube) containing 2 polyethylene catheters of different sizes (PE-160 and PE-50) connected to a pipette tip. This was introduced transvesically through an incision in the bladder dome, secured with a ligature and the pipette tip placed securely in the bladder neck. The catheter was linked to a pressure transducer and syringe pump via a 3-way stopcock to record intravesical pressure and urethral pressure and to infuse saline into the bladder. After emptying the bladder, cystometry was performed by filling the bladder with saline (0.05 mL/min).
Immunohistochemistry
Midurethral tissue (n=5/group, 20 sections/sample) was placed in 4% paraformaldehyde fixative for 16 hours, washed, and dehydrated. The tissue was embedded in paraffin, and 6-μm sections were prepared. The urethral sections were deparaffinized by incubation in Histochoice clearing agent (Amresco LLC, Solana, OH, USA) for 90 minutes, rehydrated in a graded ethanol series, washed in Tris-buffered saline (TBS; 10mM Tris-HCl, pH 7.6, 150mM NaCl), and stained with hematoxylin and eosin. IHC was performed using a labeled streptavidin biotin immunoperoxidase procedure (Universal Dako LSAB+ Kit; Dako North America Inc., Carpentaria, CA, USA). To quench endogenous peroxidase activity, the sections were treated in 3% hydrogen peroxide (Dako) for 5 minutes, washed with TBS, and incubated for 30 minutes at room temperature with monoclonal mouse anti-CAV-1, −2, and −3 antibodies (1:100; BD Biosciences, San Jose, CA, USA) diluted in TBS, followed by sequential incubations with biotinylated link (Dako) and streptavidin peroxidase (Dako) for 15 minutes each. The sections were incubated for 1 minute with substrate-chromogen solution (Dako) and counterstained for 2 minutes with hematoxylin (1:2 dilution) diluted in distilled water. The sections were then treated with 3% ammonia water (Junsei Chemical Co., Ltd., Tokyo, Japan) and mounted with Dako fluorescent mounting medium (Dako). The sections were examined and photographed using a model BX53 microscope (Olympus, Tokyo, Japan).
Western Blot
Tissue homogenates (n=10/group) were prepared from urethral tissue. Protein concentration in each sample was assayed using the BCA protein assay kit (Thermo Scientific, Rockford, IL, USA) using bovine serum albumin as the standard (Thermo Scientific), according to the manufacturer’s protocol. Equal amounts of protein (50 μg) were resuspended in sample buffer and separated by 12% sodium dodecyl sulfate-polyacrylamide gel electrophoresis. The proteins were transferred to 0.2 μm pore polyvinylidene difluoride membranes (Amersham Pharmacia Biotech, Cambridge, UK). Nonspecific binding was inhibited by 60 minutes incubation with 5% skimmed milk in TBS containing 0.05% Tween-20 (TBS-T). The membranes were incubated overnight at 4°C with monoclonal mouse anti-CAV-1, −2, or −3 antibody (1:1,000; BD Biosciences). Monoclonal mouse antibodies for CAVs (1:2,000; BD Biosciences Immunocytometry Systems, San Jose, CA, USA) and a polyclonal rabbit antibody against glyceraldehyde 3-phosphate dehydrogenase (GAPDH) (1:4,000; Cell Signaling Technology Inc., Danvers, MA, USA) were used as an internal control. The membranes were rinsed extensively with TBS-T solution and incubated for 60 minutes at room temperature with goat anti-mouse-immunoglobulin G (IgG) and goat anti-rabbit-IgG conjugated to horseradish peroxidase (1:2,500; Santa Cruz Biotechnology Inc., Santa Cruz, CA, USA) secondary antibody. The bands were visualized using the WEST-ZOL plus Western Blot Detection System (iNtRON Biotechnology Inc., Seongnam, Korea) and were imaged with the LAS-3000 system (Life-science Fujifilm Global, Tokyo, Japan). Densitometry was performed using the Multi gauge V3.0 (Fujifilm Global) chemiluminescence system and analysis software.
Statistical Analysis
Results are expressed as means±standard deviation. The urodynamic quantitative data are expressed as mean±standard error. The Mann-Whitney test was used to test the null hypothesis of no differences in mean expression levels between the groups. A P-value of <0.05 was considered significant.
RESULTS
All animals survived the 3 days after the CYP injection. Body weight between the groups was not different.
Effect of CYP-Induced Cystitis on Urodynamic Parameters
Cystometrograms were performed 3 days after the CYP injection. Nonvoiding spontaneous contractions were detected in the cystitis group. The contraction interval decreased in the cystitis group (3.9±1.0 minutes) compared to that in the control group (6.6±1.2 minutes) (P<0.05). Conversely, contraction pressure was significantly higher in the cystitis group (22.4± 0.7 mmHg) than that in the control group (11.5±0.4 mmHg) (P<0.05) (Fig. 1). The urethral pressure was decreased in the cystitis group compared to the control (4.05±2.5 mmHg vs. 5.8±2.8 mmHg, P<0.05).
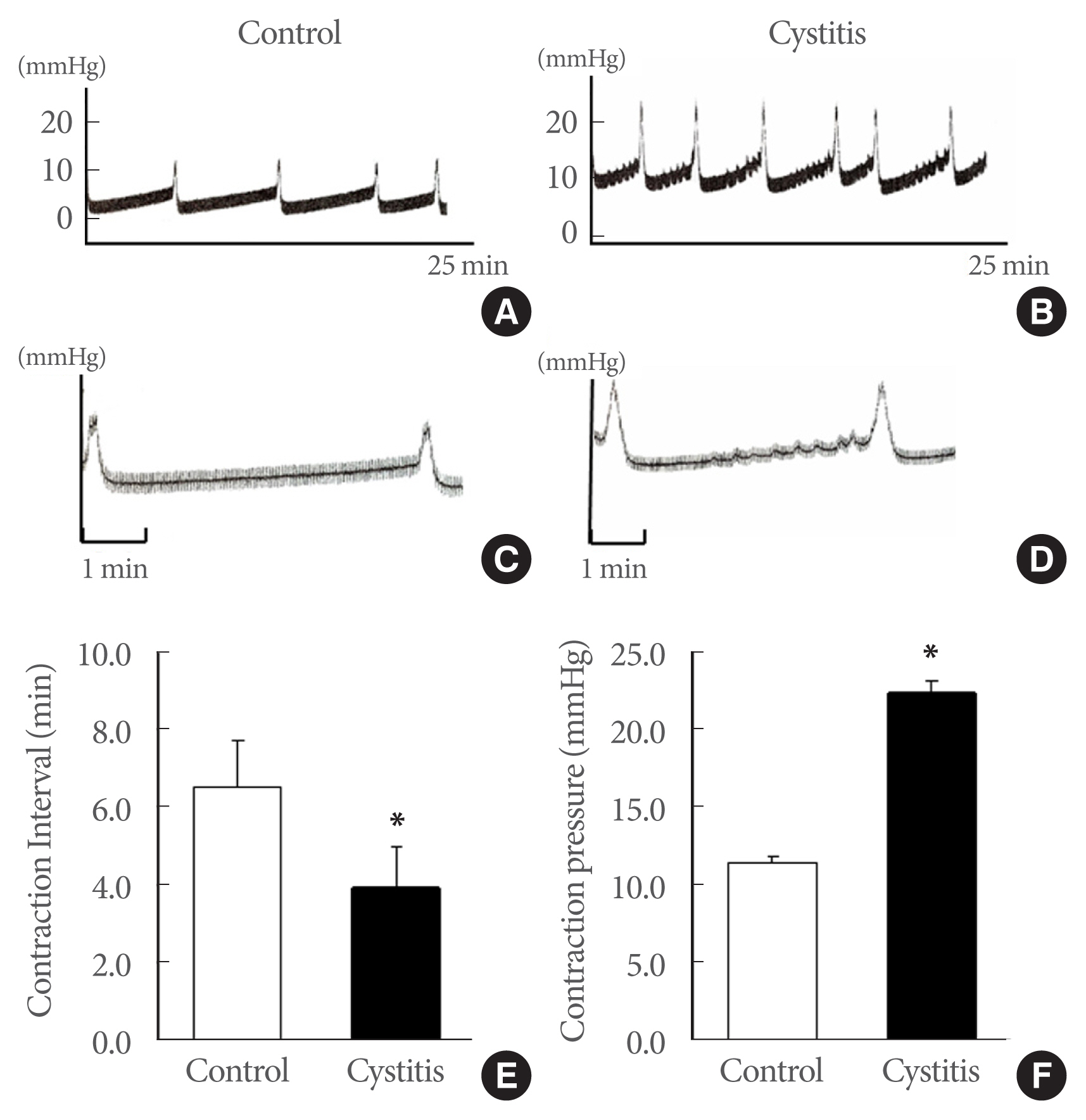
Representative urodynamic profiles: control group (A, C) and cystitis group (B, D). Nonvoiding spontaneous contractions were found in the cystitis group. The contraction interval in the cystitis group was shorter than that in the control (P<0.05). However, contraction pressure during voiding increased significantly in the cystitis compared the control group (P<0.05). The lower panels (E, F) denote the means±standard deviations of 5 experiments under each condition, as determined by cystometrograms. Panels C and D: the magnification image of each contraction of panels A and B. *P<0.05 vs. control.
Histological Changes of the Urethra
The urethral walls from inside to outside consist of urothelium, suburothelial lamina propria, inner longitudinal smooth muscle, outer circular smooth muscle, and an adventitia of loose connective tissue. The suburothelial layer is rich in microvasculature. All microvascular structures were surrounded by scattered smooth muscle bundles and connective tissue. The urethral tissue from the cystitis group showed a thickened submucosa, infiltration of inflammatory cells into the interstitium, and swelling, and irregular arrangement of muscular structure compared to those in the control group (Fig. 2).
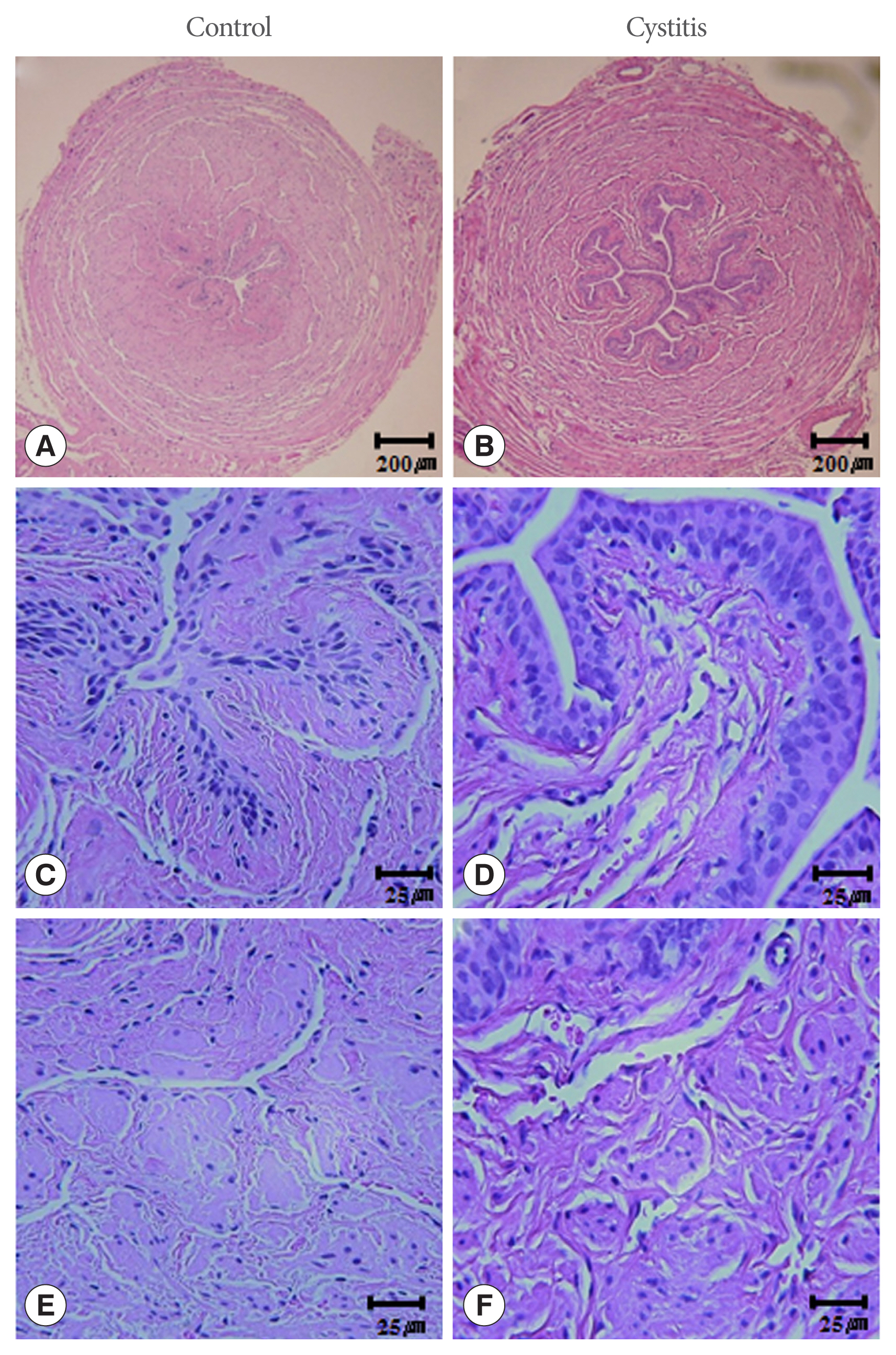
Hematoxylin and eosin staining. The urethral walls from inside to outside consist of urothelium, suburothelial lamina propria, inner longitudinal smooth muscle, outer circular smooth muscle, and an adventitia of loose connective tissue. The urethras in the cystitis group showed thickened submucosa, infiltration of inflammatory cells into the interstitium, swelling, and irregularly arranged muscle structure (B, D, and F), compared to those in the control (A, C, and E).
Urethral Expression of CAV-1, −2, and −3
Immunohistochemistry
In the urothelium and suburothelial tissue, CAV-1 and CAV-2 were expressed in the capillaries, venules, and arterioles of the suburothelial layer just beneath the urothelium, whereas CAV-3 was expressed only in urothelium. CAVs were stained brown via immunolabeling throughout the urethral tissue (Fig. 3). In the urethral smooth muscle, CAV-1 and CAV-2 were predominantly expressed in inner longitudinal smooth muscle cells, whereas CAV-3 was predominantly expressed in outer circular smooth muscle cells. The expression of CAV-1, −2, and −3 in the urethral smooth muscle was decreased in the cystitis group compared the control group (Fig. 3).

Immunohistochemistry of caveolin (CAV)-1 (A–H), CAV-2 (I–P), and CAV-3 (Q–X). CAVs were stained brown via immunolabeling (arrows). CAV-1 and CAV-2 were expressed in capillaries, venules, and arterioles in the suburothelial layer beneath the urothelium (B, F, J, and N), whereas CAV-3 was expressed in the urothelium (R and V). CAV-1, −2, and −3 were all expressed in urethral smooth muscle cells. CAV-1 and CAV-2 were mainly expressed in inner longitudinal smooth muscle cells (C, G, K, and O), whereas CAV-3 was predominantly expressed in outer circular smooth muscle cells (T, X). CAV-1, −2, and −3 expression was decreased in the cystitis group compared the control group. The horizontal scale bar at the bottom indicates magnification.
Western blot
Western blot analysis revealed CAV-1, −2, and −3 protein band at 21, 20, and 18 kDa, respectively (Fig. 4). The expression of CAV-1, −2, and −3 was significantly decreased in the cystitis group compared the control group (P<0.05) (Fig. 4).
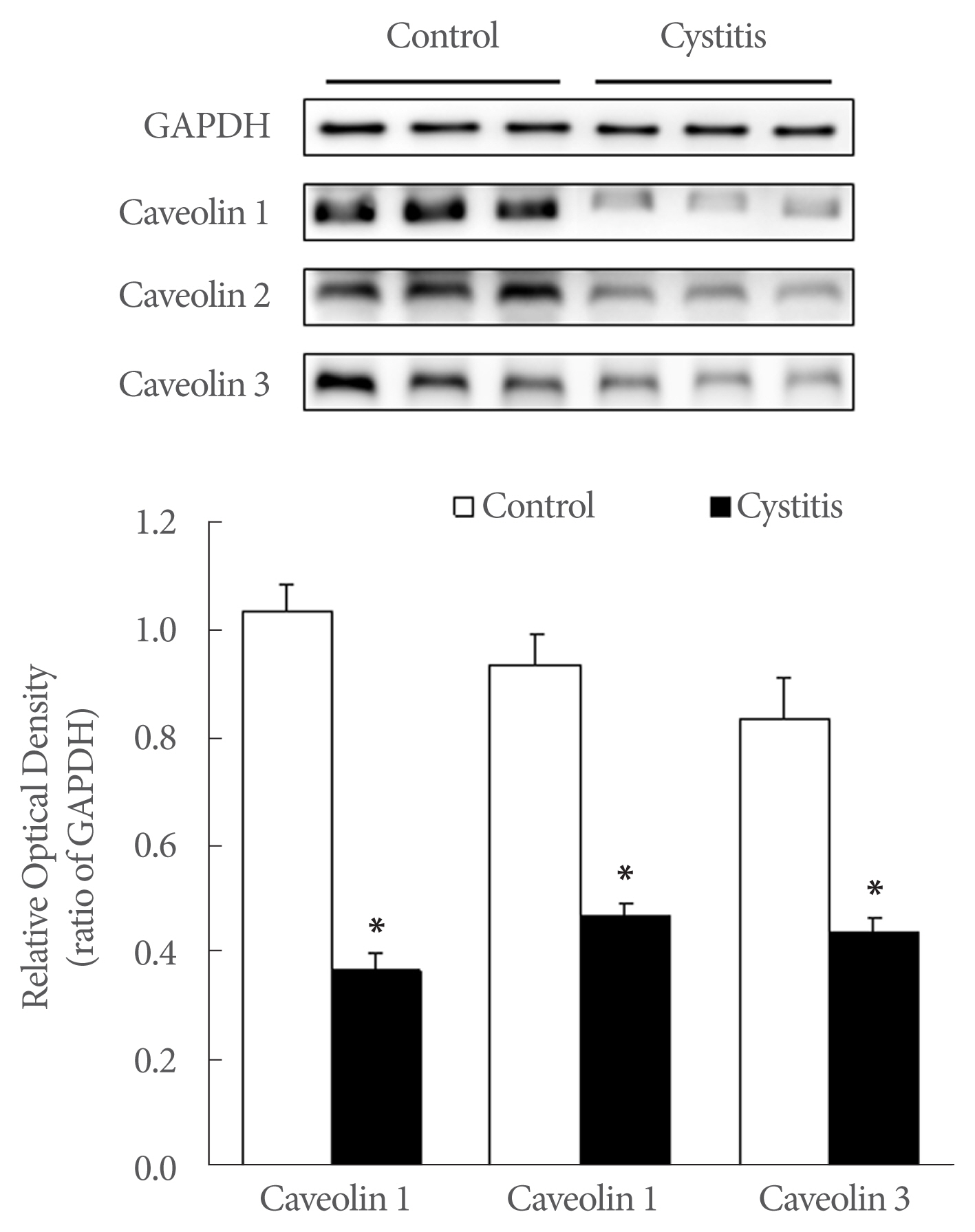
Western blot analysis of caveolin (CAV)-1, −2, and −3. The expression of CAV-1, −2, and −3 was decreased markedly in the cystitis group compared the control. The anti-CAV −1, −2, −3 antibody recognized a 21, 20, and 18 kDa band, respectively. The anti-glyceraldehyde 3-phosphate dehydrogenase (GAPDH) antibody recognized a 37 kDa band. Lower panels denote mean± standard deviation of 10 experiments for each condition, as determined by densitometry relative to GAPDH. *P<0.05.
DISCUSSION
In the present study, the expression of CAV-1, −2, and −3 in the urethra decreased significantly in the CYP-induced cystitis rat showing obvious DO of the urinary bladder. And the urethral pressure was decreased in the cystitis group compared the control. These results suggest that DO in the urinary bladder would have an impact on the urethral function and that CAV may play a role in the decreased urethral function in association with dysfunctional change of urinary bladder.
Previous studies have demonstrated that CAVs are involved in the regulation of different signaling pathways during bladder smooth muscle contraction [15–17], but little attention has been made to the expression and functional roles of CAVs in the urethra. A number of animal studies indicates that CAVs may have an important role in bladder function. Cristofaro et al. [15] concluded that CAVs differentially regulate bladder smooth muscle through stimulus-dependent potentiation or inhibition of bladder contraction before and after disruption of CAV expression. Lai et al. [16] investigated the CAVs expression of the urinary bladder by CAV-1 knockout mouse and measured smooth muscle contractions in the bladder strips evoked by electric and pharmacologic stimulation. They suggested that loss of CAV-1 is associated with disruption of M3 muscarinic subtype of cholinergic activity which resulted in impaired bladder contraction. These studies show a close functional association between CAV and signal transduction in the urinary bladder smooth muscle.
Urethral function is affected by abnormal detrusor activities [10]. Understanding the role of CAVs in a urethra with altered detrusor function may help uncovering the pathophysiological mechanism of DO or overactive bladder, and aid in developing new therapeutic targets for managing lower urinary tract symptoms such as DO.
CYP-induced chemical cystitis in animals and human studies show a mucosal erosion or edema, hemorrhagic changes, and infiltration of leukocytes in the urinary bladder [18,19]. The CYP-induced cystitis model is widely used to investigate the pathogenesis of the lower urinary tract symptoms, including overactive bladder [13]. In the present study, cystometric analysis showed a decreased contraction interval and increased contraction pressure in the cystitis group compared the control which is representing the disease model of DO. These results suggest that inflammatory change of the urinary bladder destabilizes the urinary bladder, and then these changes of the urinary bladder may affect the urethral function.
In the histopathologic study, CAV-1 and CAV-2 were expressed in the capillaries, venules, and arterioles of the suburothelial layer just beneath the urothelium in the urethra, whereas CAV-3 was expressed only in the urothelium. In the smooth muscle layers, CAV-1 and CAV-2 were predominantly expressed in the inner longitudinal smooth muscle cells, whereas CAV-3 was predominantly expressed in outer circular smooth muscle cells. The distinct expression feature suggests that CAV would have specific functional roles by each CAV subtype in the different urethral tissue layers and smooth muscle layer.
Significant advances in the understanding of bladder sensory function in terms of crosstalk between the urothelium and afferent nerves have been made in recent years. The urothelium has been thought to be a passive barrier between the urinary tract and urine. However, the urothelium is now understood to be an interactive organ that senses a variety of signals from the urinary bladder [20]. Additionally, a number of ion channels and receptors have been identified as potential targets mediating lower urinary tract function and outward signals, indicating that afferent mechanisms in the lower urinary tract may be more important and complex than previously thought [21].
This is the first study showing the possible occurrence of altered signaling process via revealing the change of the expression of CAVs in the urethra in association with bladder dysfunction. There are several potential implications of altered CAV expression on the urethra when the urinary bladder shows abnormal detrusor function. First, decreased CAV expression in the urethra may be associated with impaired urethral function induced by disruption of signal transduction pathways because CAVs have been known for the key regulators of signaling process involved in cellular reactions and smooth muscle contractions. The urothelium can sense chemical and mechanical stimuli that may relay the status of the urothelial environment to the underlying nervous and muscular systems [22]. CAVs seem to be involved in transmitting signal transducing activity from the urothelium to afferent nerves and then to the smooth muscle. Second, the expressional localization of CAVs in the urethra may be associated with the intrinsic change of the urethra that occur with functional changes of the urinary bladder such as DO. The submucosal layer is highly vascularized and likely to play a role in maintaining of urethral coaptation and keeping the urethra closed [23]. Decreased expression of CAV-1 and CAV-2, which are mainly localized in the microvasculature, could activate endothelial NOS and produce high nitric oxide levels [24,25], which would induce vasodilatation and change blood supply and urethral physiology. The difference in localization among CAV-1, −2, and −3 in the urethra suggests that these molecules have distinct functional roles by CAV subtype in specific urethra muscle layers. The decreased expression of CAVs in urethral smooth muscle is likely to be involved in the decreased urethra contractility, as decreased expression of CAVs may affect the intracellular calcium which regulates smooth muscle tone. Third, direct inflammatory changes in the urethra from CYP-induced cystitis would be one of the reasons of the altered CAV expressions in the urethral tissues.
A limitation of our study is that the precise pathophysiological mechanism or functional activity of CAVs on the urethra was not fully unveiled. Further studies are needed to understand the exact roles of CAVs in the urethra by revealing their functional responses assessed by in vivo and in vitro study involved in other pharmacological agents mediating the signal transduction of the urinary bladder and urethra. More information about the roles of CAVs may be obtained using a CAV deficiency model. Additionally, urethral cell culture and electron microscopic studies on urethral CAVs will contribute to a further understanding of their functional roles in the urethra. Taken together, our results strongly suggest that DO may significantly downregulate the expression of CAV in the urethra, providing presumptive evidence that CAVs are involved in the functional change of the urethra in association with DO, probably by modifying afferent signaling pathways related with CAVs.
In conclusion, our study indicates that the functional change of urethra and the decreased expression of urethral CAV-1, −2, and −3 was observed in the animal model of DO by CYP-induced cystitis rat. These data suggest that urethral CAVs may, in part, be involved with the functional change of urethra in association with DO of urinary bladder.
Notes
Fund/Grant Support
This study was supported by a grant of the Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education, Science and Technology (2014-1825); by a grant CRI 17015-1, Chonnam National University Hospital Research Institute of Clinical Medicine.
Research Ethics
All experiments were approved by the Ethics Committee of Chonnam National University Medical School (CNU IACUC-H-2015-10).
Conflict of Interest
No potential conflict of interest relevant to this article was reported.
AUTHOR CONTRIBUTION STATEMENT
• Conceptualization: SOK
• Data curation: HJC, HYJ
• Formal analysis: HJC, HYJ
• Funding acquisition: SOK
• Project administration: SOK
• Writing - original draft: HJC, HYJ
• Writing - review & editing: SOK