• HIGHLIGHTS
- Ten-eleven translocation methylcytosine dioxygenase 1 (TET1) acts as a repressor for α-synuclein gene (SNCA).
- Postmortem human Parkinson disease samples express low levels of TET1 compared to the control samples.
INTRODUCTION
Parkinson disease (PD) is a progressive neurological disorder characterized by the loss of dopaminergic neurons in the substantia nigra (SN) and accumulation of α-synuclein (α-SYN) [1,2]. The SNCA gene encodes α-SYN protein and can over-produce α-SYN, which accumulates abnormally in the brains of people with PD [3]. Excessive expression of α-SYN is strongly associated with familial and sporadic PD [4-7]. Moreover, many studies related to SNCA-intron1 hypomethylation, transcription factors, polymorphisms in the distal enhancer-binding region, microRNA-mediated regulation, and upstream dinucleotide repeat polymorphisms (NACP-Rep1) have powerfully demonstrated the mistranslation and epigenetic regulation of SNCA in PD [8-15]. SNCA harbors a strong CpG island (CGI) in the upstream regulatory region encompassing promoter and intron1 [16]. Our previous study has shown that the CpGs in the intron1 region remain equally hypomethylated in control and PD subjects, indicating a limited role of DNA methylation in the regulation of SNCA in PD [17]. Other studies have also observed similar to our findings [18-20]. However, this hypomethylated CGI of SNCA and associated histone modifications might play roles in recruiting other epigenetic modifiers and transcription factors to regulate α-SYN expression in dopaminergic neurons.
The ten-eleven translocation (TET) protein family includes 3 members (TET1-3), and all 3 TET proteins can subsequently oxidize the methyl group of 5-methylcytosine (5mC) to yield 3 different forms of 5-hydroxymethylcytosine (5hmC), 5-formylcytosine, and 5-carboxylcytosine [21,22]. The ten-eleven translocation methylcytosine dioxygenase 1 (TET1), a member of the TET family of enzymes, is one of the CGI binding proteins and is known for its role in catalytic conversion of 5mC to 5hmC [23]. The discovery of TET1 partially contributed to solving the puzzle of DNA [23-25]. Interestingly, TET1 has been shown to work as a dual function oxidase in mouse embryonic stem cells, allowing repression and activation of 2 different target genes [26]. It can also recruit other corepressor molecules, Sin3A, which can suppress gene transcription [27]. TET1 preferentially binds to an unmethylated CGI-containing promoter through its CXXC domain and regulates gene expression [28-30]. This protein has also been shown to bind the bivalent promoters, which are marked by both histone 3 lysine 4 trimethylation (H3K4me3; activation mark) and histone 3 lysine 27 trimethylation (H3K27me3; repression mark) modifications [26,30,31].
This research investigates whether TET1 binds to SNCA-intron1 and regulates gene expression. Since SNCA is generally a bivalent promoter and remains hypomethylated in the neuronal cells, we sought to examine whether the regulatory function of TET1 is independent of its catalytic activity. Our study aims to unravel a novel epigenetic mechanism by which α-SYN expression is controlled in PD.
MATERIALS AND METHODS
Postmortem Brain Tissues
We used postmortem brain samples from the SN region (control [n=8], PD [n=17]). All human postmortem brain samples were procured from NIH Neurobiobank and were used in accordance with the principles of the Declaration of Helsinki of the World Medical Association. All the samples were ethnicity, age, and sex-matched. The demographic data of all participating samples are presented in Supplementary Table 1.
Cell Culture
ReNcell VM cell line derived from human fetal ventral mesencephalon was obtained from EMD Millipore (SCC008). The cells were cultured following the previously described [32]. ReNcell VM was maintained in a medium containing DMEM/F12 (Thermo Fisher Scientific Inc., Waltham, MA, USA; SH 30023FS) supplemented with B27 supplement (Life technologies, Carlsbad, CA, USA; 17504-044), Glutamax (Life technologies, 35050-061), 10-units/mL heparin sodium salt (Tocris Bioscience, Ellisville, MO, USA; 2812), 50-μg/mL Gentamicin (Life technologies, 15750-060), 20-ng/mL epidermal growth factor (Millipore, Burlington, MA, USA; GF144), and 20-ng/mL basic fibroblast growth factor (PeproTech, Seoul, Korea; AF-100-18B) on laminin-coated tissue culture dishes. Differentiation of the cells was accomplished by changing the medium mentioned above without growth factors for 14 days.
HEK293T cells were cultured in DMEM/high glucose medium (Thermo Fisher Scientific, SH30243FS) supplemented with 10% fetal bovine serum (Atlanta Biological Pvt. Ltd., Kolkata, India; S10350H). All cells were maintained in a humidified atmosphere with 5% CO2 at 37°C.
DNA Constructs and Transfection
The CXXC domain (amino acid 528-674) of TET1 was amplified from cDNA of undifferentiated ReNcell VM. The 441 bp amplicon containing EcoRI/XhoI restriction enzyme sites was cloned into pAAV-IRES-hrGFP vector harboring 3X flag tag (Clontech Laboratories Inc., Mountain View, CA, USA). To generate the ReNcell VM stably overexpressing the TET1-CXXC domain, the cDNA encoding TET1-CXXC-flag from pAAV-IRES-hrGFP vector was subcloned into EcoRI/ApaI sites of pLVX-DsRed-Monomer-N1 vector (Clontech Laboratories Inc.). The catalytic domain of TET1 (amino acids 1418-2136) and its inactive catalytic form cloned in pAAV-EF1a-HA-hTET1CD-WPRE-PolyA vector were a kind gift from Hongjun Song (Addgene plasmids # 39454 and 39455). According to the manufacturer’s instructions, ReNcell VM or HEK293T cells were transfected with plasmid DNA using X-fect transfection reagent (Clontech Laboratories Inc., PT5003-2).
Generation of Stable Cell Line
ReNcell VM stably expressing the TET1-CXXC domain were generated by lentiviral transduction. The pLVX-TET1-CXXC-DsRed-Monomer-N1 vector was cotransfected with lentiviral packaging plasmids pLP1, pLP2, and pLP/VSVG (Life Technologies, K4944-00) at a 1:1:1:1 ratio using X-fect transfection reagent in HEK293T cells. 48 hours after transfection, the lentiviral particles containing the medium were collected and centrifuged briefly at 500 g for 10 minutes to remove the remaining cell debris. Lentiviral particles were treated to ReNcell VM grown on 24 well plates. Following 48-hour treatment with lentiviral particles, the cells were positively selected under antibiotic puromycin (2 mg/mL) (Acros Organics, Veneto, Italy; 227420100).
Knockdown of TET1
Knocking down of the human TET1 gene was achieved by either lentiviral particles containing short hairpin RNA (shRNA) constructs or by small interfering RNA (siRNA), as mentioned in the text. The shRNA plasmid cocktails were purchased from Open Biosystems (RHS4531; now acquired by Horizon Discovery, Cambridge, UK). The shRNAs were embedded in miR30a backbone in pGIPZ vector; the sequences are shown in Supplementary Table 2. As mentioned above, lentiviral particles were made, and stable shRNA cell lines (shTET1) were made with individual shRNA following antibiotics selection. For knockdown assays, TET1-shRNA stable line and control (a nontargeting shRNAmiR-pGIPZ) stable line were differentiated for 14 days before harvesting them for western blot analysis. These cell lines stably express TET1 knockdown sequences. No abnormalities were observed in the differentiation of these shRNA stable lines compared to the control (empty vector and wildtype) lines. Additionally, we carried out knockdown of TET1 using transient overexpression of validated siRNAs in wild-type ReNcell VM. Double-stranded siRNAs targeting TET1 (siTET1) and nontargeting siRNA (scrambled siRNA for a negative control) were obtained from Thermo Fisher Scientific (s37192). The sequences for siTET1 were S ense 5´-GAAGCUGAAAAAUCGAAAAtt-3´, Antisense 5´-UUUUCGAUUUUUCAGCUUCtg-3´. The ReNcell VM was transfected with siTET1 or scrambled siRNA using Lipofectamine RNAiMAX (Thermo Fisher Scientific, 13778030) as the manufacturer’s instructions recommend.
Chromatin Immunoprecipitation
Chromatin immunoprecipitation (ChIP) was performed following protocol for the EZ ChIP kit (Millipore, 17-371) with the following modifications as described in Guhathakurta et al. [33]. The shearing of chromatin was standardized, as shown in Supplementary Fig. 1. Supplementary Table 3 shows a list of antibodies used in ChIP.
RT-PCR and Real Time-qPCR
Total RNA was extracted from the ReNcell VM using Trizol reagent (Invitrogen, Waltham, MA, USA). cDNA was synthesized as described in the manufacturer’s protocol (GenDEPOT, Katy, TX, USA; R5600-050). The sequences of specific primers are shown in Supplementary Table 4. Relative TET1 mRNA levels were determined by real time-quantitative PCR using Power SYBR Green Master Mix (Thermo Fisher Scientific, 4367659). Real time-qPCR reaction programs were 50°C for 2 minutes, 95°C for 2 minutes, and 40 cycles of 95°C for 3 seconds and 60°C for 30 seconds. The relative mRNA level was normalized to GAPDH expression and calculated by the 2-ΔΔCt method.
Western Blot
Proteins (15–75 μg/sample) were loaded on 5 or 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis and transferred to a polyvinylidene fluoride membrane. The membranes were blocked by 5% nonfat milk in Tris-buffered saline containing 0.05 % Tween-20 (TBS-T) for 1 hour at room temperature (RT) and incubated with respective primary antibodies at 4°C overnight. And then, the membrane was incubated with respective secondary antibodies for 1 hour at RT. The protein bands were visualized by Enhanced Chemiluminescent detection Reagents (Thermo Fisher Scientific, 45-010-090). A list of antibodies used in western blot is presented in Supplementary Table 3.
Dot-Blot
DNA Dot-blot for detecting 5hmC was performed following the protocol outlined by Ito et al. [22] 2011 with modifications. Genomic DNA was isolated from the untransfected and transfected ReNcell VM with the TET1-catalytic domain or its inactive form. Three different concentrations with the same volume (125 ng, 250 ng, and 500 ng) of each sample were blotted on Zeta-Probe GT Genomic Tested Blotting Membranes (Bio-Rad Laboratories, Inc., Hercules, CA, USA). The dots were UV cross-linked (UVC 1000) on the membrane and blocked in 5% nonfat milk in TBS-T for 1 hour at RT. The membrane was incubated with anti-5hmC antibody (1:10,000; Active Motif, Carlsbad, CA, USA, 39769) at 4°C overnight. Then, the membranes were incubated with a secondary antibody for 1 hour at RT. The membrane was visualized with Super Signal West Pico Chemiluminescent Substrates (Thermo Fisher Scientific, 34077).
Statistical Analyses
Data are presented as mean ±standard error of the mean. Statistical analyses were conducted using GraphPad Prism v.9.3.1 (GraphPad Software Inc., San Diego, CA, USA). An unpaired Student t-test for cell line data and a Mann-Whitney t-test for human subjects were performed to compare the mean between the 2 groups. The data for methylation was prepared using Quantification Tool for Methylation Analysis (QUMA) software (http://quma.cdb.riken.jp). The level of statistical significance was considered if the P-value was <0.05.
RESULTS
SNCA Regulatory Region Shows Bivalent Histone Marks
We first investigated the epigenetic structure of SNCA in the SN from adult human brain samples in NIH Roadmap epigenomics data. It showed both H3K4me3 and H3K27me3 marks in the upstream regulatory region of SNCA. There was a sharp peak of H3K4me3 encompassing the transcription start site (TSS) and CpG island region. The H3K27me3 was distributed throughout the gene body (Fig. 1A). We also checked 2 histone marks in SNCA-intron1 with undifferentiated/differentiated human neuronal cells. Undifferentiated/differentiated ReNcell VM showed enrichment of both H3K4me3 and H3K27me3. Differentiated ReNcell VM demonstrated stronger co-presence than undifferentiated ReNcell VM (Fig. 1B). The methylation states of SNCA-intron1 of undifferentiated and differentiated ReNcell VM exhibited largely unmethylated CGI (Fig. 1C, D), similar to our previously reported human postmortem brain samples [17].
TET1 Binds to the Human SNCA-Intron1 Region and Represses α-SYN Expression
It has been reported that TET1 binds to the unmethylated CGI-containing monovalent promoters (marked by H3K4me3 only) or bivalent promoters (marked by both H3K4me3 and H3K27me3) [28]. TET1 is a large protein of 2,136 amino acids with a molecular weight of 232 kDa [23]. This protein has 2 distinct domains with different functions, a cysteine-rich zinc finger CXXC domain for DNA binding and a catalytic domain for 5mC hydroxymethylation (Fig. 2A). The hypomethylated CGI of SNCA-intron1is bivalently marked, implying an ideal region where TET1 may bind. We first verified the expression of TET1 and found abundant protein levels in undifferentiated and differentiated ReNcell VM. The binding of TET1 at the SNCA-intron1 was examined in human postmortem SN tissues using the ChIP assay. As expected, we observed TET1 bound to the SNCA-intron1 in human SN tissues (Fig. 2B). Since TET1 is a relatively large protein, overexpressing TET1 was challenging. We tried to overexpress the full-length TET1; however, we remained unsuccessful despite our repeated attempts to adhere to different transfection/electroporation protocols (Supplementary Fig. 2). To circumvent the situation, we subcloned and overexpressed the DNA binding CXXC domain and hydroxymethylating catalytic domains separately to see their effect on α-SYN expression.
Flag-tagged DNA binding TET1-CXXC domain was overexpressed in HEK293T cells to determine its binding to this region. We found that the TET1-CXXC domain significantly binds to the SNCA-intron1 (t=-2.927, P=0.021) (Fig. 2C). To confirm how the binding of TET1 affects α-SYN expression in neuronal cells, we established the ReNcell VM cell line stably expressing the TET1-CXXC domain only. Overexpression of the TET1-CXXC domain showed a significant decrease in α-SYN levels compared to cells expressing the empty vector (t=3.456, P=0.013) (Fig. 2D). We investigated whether knockdown of TET1 using either siRNA or shRNAs could affect the α-SYN levels (Fig. 2E). TET1 siRNA or shRNA (shRNA#11) significantly decreased the TET1 levels compared to the scramble siRNA (t=2.566, P=0.026) or compared to the control lines (t=3.077, P=0.020). On the other hand, TET1 knockdown significantly increased α-SYN levels (mRNA: t= -3.431, P=0.007; Protein: t=-3.742, P=0.006).
TET1-Catalytic Domain Plays a Limited Role in Regulating α-SYN Expression
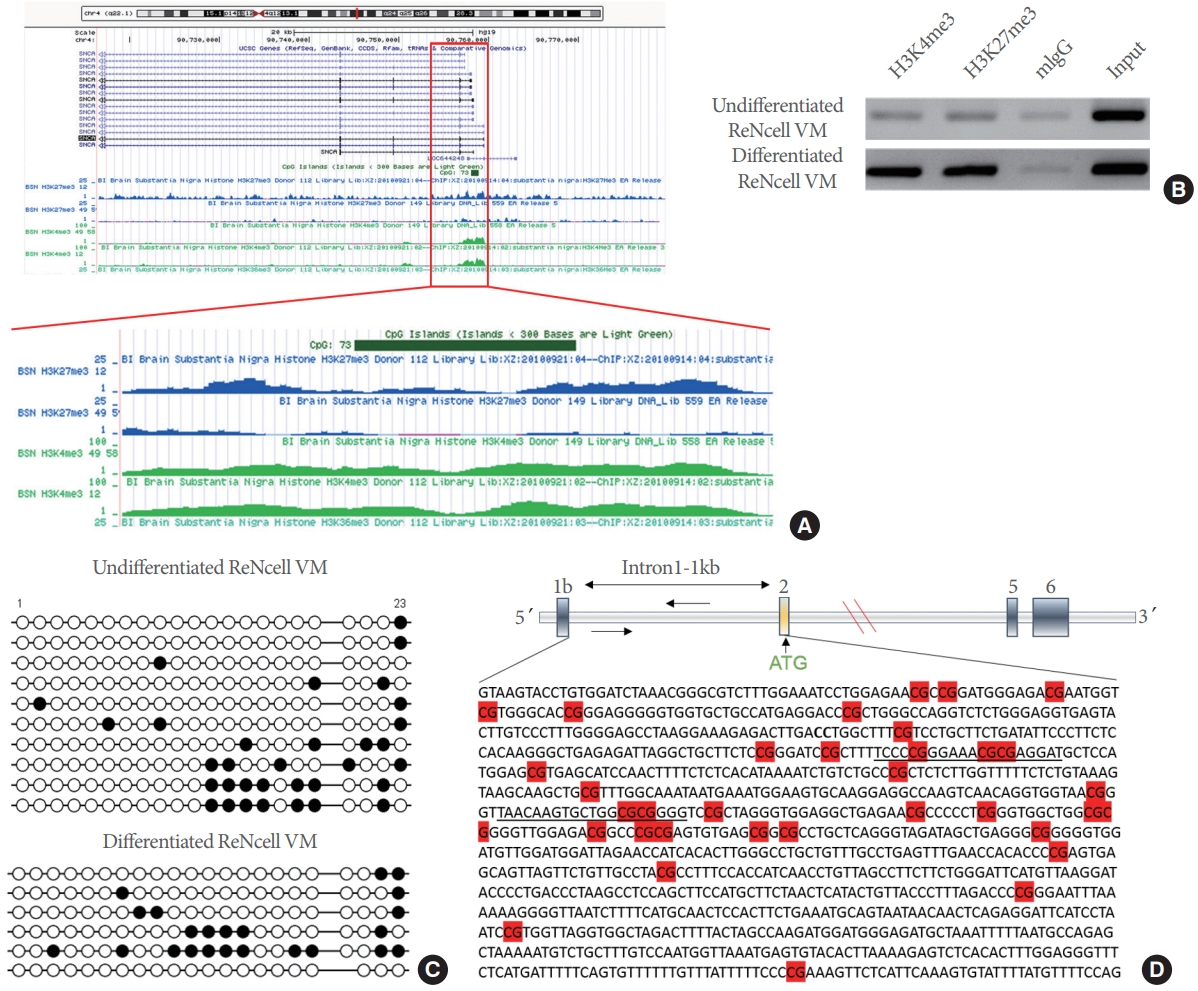
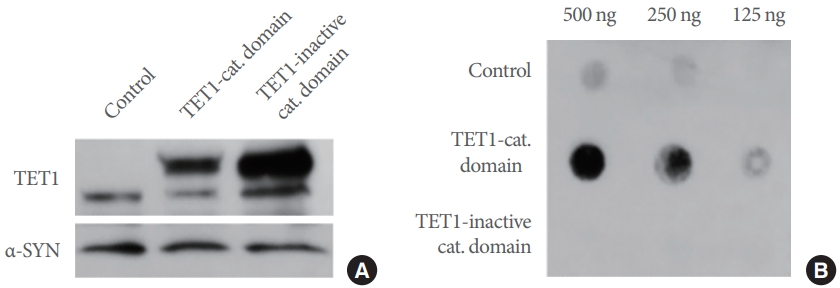
We showed that TET1 catalyzes 5mC to 5hmC conversion using its C-terminal catalytic domain in Fig. 2A. Therefore, we examined whether the catalytic domain affects α-SYN expression. Overexpression of the TET1-catalytic domain or the catalytically inactive domain in undifferentiated ReNcell VM did not alter α-SYN levels, suggesting that TET1-mediated α-SYN repression is independent of its catalytic activity (Fig. 3A). We confirmed that overexpression of the catalytic domain increased endogenous 5hmC levels in the cells in contrast to its inactive catalytic domain (Fig. 3B).
PD Cases Express Low Levels of TET1
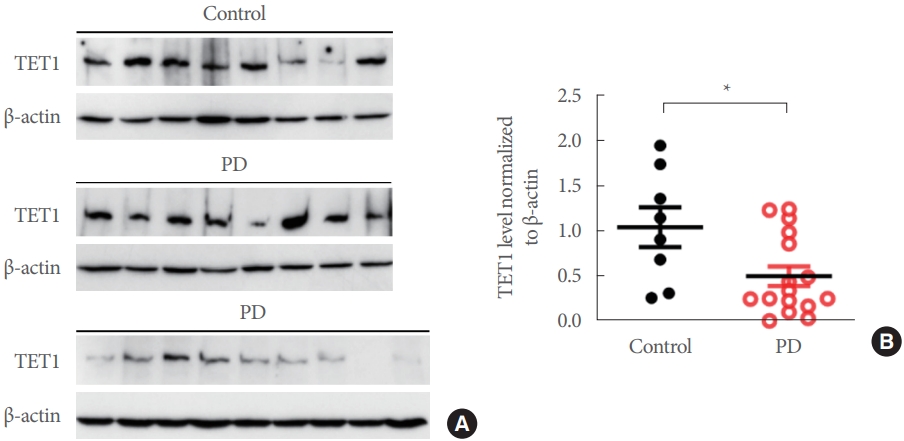
Next, we examined the levels of TET1 in postmortem human SN brain samples. Interestingly, we found that the TET1 levels of PD subjects were significantly less than the control subjects (Mann-Whitney t-test, P<0.05) (Fig. 4A, B). In our recently published article, we showed that the same postmortem brain samples of PD cases had significantly higher levels of α-SYN as compared to the controls [33].
DISCUSSION
This study showed a novel mechanism for regulating α-SYN expression by TET1 in neuronal cells and postmortem brain samples. Our study is the first to describe the importance of TET1 in α-SYN regulation in PD pathogenesis. Epigenetic regulation of SNCA concerning PD has only been focused on the methylation structure of this gene [3].
TET1 is known to catalyze DNA hydroxymethylation and has been shown to act as both activator and repressor for gene expression [26]. The stable expression of 5hmC in the adult brain suggests a specific role of TET1 in the nervous system [24]. It has also been shown that TET1 plays a crucial role in active demethylation in the adult mouse brain [35]. As mentioned earlier that TET1 binds preferentially to the unmethylated CGI-containing promoters, it might have roles other than DNA hydroxymethylation in the case of hypomethylated genes. The regulatory region of human SNCA comprises a much larger CGI with significant hypomethylation compared to the murine genome [3]. As TET1 binds to large unmethylated CGIs, it can be expected that the effect of TET1-mediated regulation of SNCA in humans could be more pronounced than in murine systems. Our previous study showed that the SNCA-intron1 region remains significantly hypomethylated in the postmortem SN tissues from both PD and matched control subjects without any bias for the disease [17]. We observed that TET1 binds to this SNCA-intron1 regulatory region encompassing a hypomethylated CGI in human SN. We also found that this region of SNCA has both H3K4me3 and H3K27me3 in the differentiated neuronal cell line. Since TET1 can bind to bivalent promoter-containing genes, neuronal SNCA is an ideal target. Moreover, the region near the gene’s TSS also contains a putative TET1 binding sequence (5´-GCGCCGC-3´) [26].
Interestingly, we found that the CXXC-DNA binding domain of TET1 binds to this region and is enough to repress the gene expression without contribution from the catalytic domain, which is responsible for DNA hydroxymethylation. This result can be explained by the fact that CGI of the regulatory region of SNCA remains hypomethylated in the neuronal cells. At the same time, the knockdown of TET1 in these cells led to a significant increase in α-SYN levels, which indicates that TET1 has a repressive effect on SNCA expression. Therefore, hydroxymethylation and subsequent demethylation-mediated regulation of SNCA is less likely to happen even in the TET1-catalytic domain overexpressed condition. However, the global hydroxymethylation in the cell was significantly increased upon overexpression of the TET1-catalytic domain.
Although the molecular mechanism of how the TET1-CXXC domain alone can inhibit gene expression is unknown, the CXXC domain might recruit other corepressors and repress the transcription altogether. We have also seen that overall TET1 levels in PD patients are significantly lower than in the controls. Our group has recently shown that occupancy of H3K4me3, a transcription initiation/favoring histone posttranslational modification, at the SNCA-promoter/intron1 region is significantly higher in PD patients, in turn, may make it less favorable for TET1 binding in the PD patients selectively [33].
Deregulated expression and aggregation of α-SYN are a long-standing observation in PD. However, the mechanism remains elusive. This report provided molecular and epigenetic insight into this gene’s deregulation in disease conditions. Although TET1 is known for its hydroxymethylating activity, we observed that the DNA binding domain of TET1 could repress SNCA expression by physically binding the intron1 region of the gene. This study links to how the epigenetic niche of SNCA is altered in PD specifically, which could be a target for therapeutic intervention for the disease.