INTRODUCTION
Transient cerebral ischemia is induced by temporary deprivation of blood flow to the brain, and results in neuronal degeneration. Pyramidal neurons in the CA1 region are the most vulnerable cells to ischemic-reperfusion injury, and loss of hippocampal CA1 neurons causes long-term learning deficits [1].
Astrocytes are activated (reactive) in response to stroke, trauma, tumor growth, or neurodegenerative diseases. Astrocytic activation results in so-called "reactive astrogliosis", which is an inflammatory response characterized by astrocytic proliferation and hypertrophy following brain injury [2]. Glial fibriallary acidic protein (GFAP) is the main constituent of intermediate astrocytic filaments. GFAP expression has been shown to increase after transient ischemic stroke in rats [3]. Moreover, alterations in GFAP immunoreactivity are associated with neuronal cell death in the ischemic CA1 region [4], and GFAP expression has been said to represent a hallmark of reactive astrogliosis [5]. Activated microglia readily undergo dramatic changes in morphology and the surface expression of molecules such as major histocompatibility complex, which is recognized by the antibody to CD11b. CD11b is located on the plasma membrane of microglia, and CD11b expression is increased during the microglial activation [6]. 1-Methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) has been used in animal models of Parkinson disease, and MPTP administration causes enhanced CD11b expression in the striatum and substantia nigra [7].
The phosphoinositide 3-kinase (PI3K)/protein kinase B (Akt) signaling pathway plays a central role in intracellular processes related to cell survival and proliferation. PI3K generates phosphatidylinositol triphosphate, which activates the serine/threonine kinase Akt. Akt then modulates cellular activation, the inflammatory response, and apoptosis [8]. Thus, it is not surprising that decreased Akt activity is involved in ischemia-induced cell death [9], while increased Akt activity is associated with neuroprotective effects in ischemic gerbils [10].
Apoptosis maintains homeostasis; however, inappropriate or excessive apoptosis has been implicated in several types of neurodegenerative disorders, including ischemia [7,11,12]. Apoptosis after cerebral ischemia is one of the major pathways that lead to the process of cell death [13]. Terminal deoxynucleotidyl transferase-mediated dUTP nick end labeling (TUNEL) staining is an assay that detects DNA fragmentation, a characteristic of apoptotic cell death [11]. One important characteristic of apoptosis is the activation of caspases. Caspase-3 is one of the most widely studied members of the caspase family, and is one of the key initiators of apoptosis [14]. In addition, Bax and Bcl-2 are major proteins that have been shown to regulate apoptosis. Specifically, Bcl-2 is an antiapoptotic protein whose expression protects against cell death, while Bax is a proapoptotic protein that it is expressed abundantly during apoptosis. Increasing the ratio of Bax to Bcl-2 has commonly been used to determine the induction of apoptosis in several tissues [12,15].
Alkaloids are found in bacteria, fungi, plants and animals, although their distribution within each kingdom is quite limited. Despite their structural diversity, alkaloids share many physical and chemical properties. Berberine is a type of isoquinoline alkaloid extracted from Berberis vulgaris L, and is used to treat various diseases, including tumors, diabetes mellitus, cardiovascular deregulation, immunology disorders, neurodegeneration, and neuropsychiatric disorders [16,17]. The neuroprotective effect of berberine against cerebral ischemia has been reported [18,19]; however, the effect of berberine on apoptosis under ischemic conditions is not clear.
In the present study, we investigated the effects of berberine on global ischemia-induced apoptosis focusing on the PI3K/Akt signaling pathway in the gerbil hippocampus. For this, a step-down avoidance task was employed to assess short-term memory, the TUNEL assay was used to evaluate DNA fragmentation, immunohistochemistry for GFAP, CD11b, and caspase-3, and western blot for the identification of PI3K, Akt, Bax, Bcl-2, and cytochrome c levels.
MATERIALS AND METHODS
Experimental Animals
Adult male Mongolian gerbils (13 weeks old) were used in this experiment. All experimental procedures were performed in accordance with the animal care guidelines of the National Institutes of Health and the Korean Academy of Medical Sciences. Gerbils were housed under controlled temperature (20Ōäā┬▒2Ōäā) and lighting (7 AM to 7 PM) conditions with food and water available ad libitum. The gerbils were randomly divided into five groups (n=10 in each group): sham-operation group, ischemia-induction group, ischemia-induction and 20 mg/kg berberine-treated group, ischemia-induction and 50 mg/kg berberine group, and ischemia-induction and 80 mg/kg berberine group. Gerbils in berberine-treated groups received berberine (Sigma Chemical Co., St. Louis, MO, USA) orally once a day for 14 consecutive days, starting one day after surgery. Gerbils in sham-operation and ischemia-induction groups received an equal amount of distilled water for the same duration.
Induction of Transient Global Ischemia
Transient global ischemia was induced as previously described [11]. Briefly, gerbils were anesthetized with Zoletil 50 (10 mg/kg, intraperitoneally; Vibac Laboratories, Carros, France), and following bilateral neck incisions, both common carotid arteries were exposed and occluded with aneurysm clips for 7 minutes. Clips were then removed to restore cerebral blood flow. Body and rectal temperature was maintained at 36Ōäā┬▒0.5Ōäā during surgery using a Homeothermic Blanket Control Unit (Harvard Apparatus, Massachusetts, MA, USA) that enveloped the body and head. After recovery, animals were monitored for an additional 2 hours to prevent hypothermia. Animals in the sham-operation group were treated identically, except that the common carotid arteries were not occluded after neck incisions.
Step-Down Avoidance Task
In order to evaluate short-term memory, latency in the step-down avoidance task was determined, as per the previously described method [20]. Fourteen days after surgery, gerbils were trained in a step-down avoidance task. Rats were positioned on a 7├Ś25-cm platform with a height of 2.5 cm, and then allowed to rest on the platform for 2 minutes. The platform faced a 42├Ś25-cm grid of parallel 0.1-cm caliber stainless steel bars, which were spaced 1 cm apart. In the training sessions, animals received a 0.5-mA scramble foot shock for 3 seconds immediately upon stepping down. Retention time was assessed 15 days after surgery. The interval for rats stepping down and placing all four paws on the grid was defined as the latency. A latency over 300 seconds was counted as 300 seconds.
Tissue Preparation
For brain tissue preparation, animals were fully anesthetized with Zoletil 50, transcardially perfused with 50mM phosphate-buffered saline (PBS), and fixed with freshly prepared solution consisting of 4% paraformaldehyde in 100mM phosphate buffer (PB, pH 7.4). Brains were then removed, postfixed in the same fixative overnight, and transferred into a 30% sucrose solution for cryoprotection. Coronal sections of 40-┬Ąm thickness were made using a freezing microtome (Leica, Nussloch, Germany).
Immunohistochemistry
Immunohistochemistry was performed according to the previously described method [21,22]. Free-floating tissue sections were first incubated in 3% H2O2 for 30 minutes to block endogenous peroxidase activity. Sections were then incubated in blocking solution (1% bovine serum albumin and 10% horse serum in 0.05M PBS) for 2 hours at room temperature, and were incubated overnight with antimouse GFAP antibody (1:1,000; Cell Signaling Technology, Beverly, MA, USA), antirat CD11b antibody (1:500; Serotec, Raleigh, NC, USA), and antimouse caspase-3 (1:500, Santa Cruz Biotechnology, Dallas, TX, USA) at 4Ōäā. The next day, sections were incubated for 1 hour with biotinylated secondary antibody (1:200; Vector Laboratories, Burlingame, CA, USA) at room temperature, and were subsequently incubated with a Vector Elite ABC kit (Vector Laboratories) for 1 hour at room temperature. The antibody-biotin-avidin-peroxidase complex was visualized using 0.02% 3,3'-diaminobenzidine tetrahydrochloride (DAB) and 0.03% H2O2 in 50mM Tris-buffer (pH 7.6) for approximately 5 minutes and was then washed with PBS and mounted onto gelatin-coated slides. Sections were mounted onto gelatinized glass slides, air dried, and cover slides were mounted using Permount (Thermo Fisher Scientific Inc., Fair Lawn, NJ, USA).
TUNEL Staining
To visualize DNA fragmentation, TUNEL staining was performed using an In Situ Cell Death Detection Kit (Roche, Mannheim, Germany), according to a previously described method [23]. Sections were postfixed in ethanol-acetic acid (2:1) and rinsed. The sections were then incubated with proteinase K (100 ┬Ąg/mL), rinsed, and incubated in 3% H2O2, permeabilized with 0.5% Triton X-100, rinsed again, and incubated in the TUNEL reaction mixture. The sections were rinsed and visualized using Converter-POD with 0.03% DAB. Mayer's hematoxylin (DAKO, Glostrup, Denmark) was used as a counterstain, and sections were mounted onto gelatin-coated slides. Slides were then air-dried overnight at room temperature, and coverslips were mounted using Permount (Thermo Fisher Scientific Inc.).
Western Blot Analysis
Western blot was performed according to previously described methods [21]. Tissue samples harvested from the hippocampus were lysed in protein lysis buffer containing 50mM Tris-HCI (pH 7.5), 150mM NaCl, 0.5% deoxycholic acid, 1% nonidet-P40 (NP40), 0.1% sodium dodecyl sulfate (SDS), 1mM phenylmethylsulfonyl fluoride, and 100 ┬Ąm/mL leupeptin. Protein concentration was measured using a colorimetric protein assay kit (Bio-Rad, Hercules, CA, USA). Proteins of 40 ┬Ąg were separated on SDS-polyacrylamide gels and transferred onto a nitrocellulose membrane (Schleicher & Schuell GmbH, Dassel, Germany). The membranes were incubated with 5% skim milk in Tris-buffered saline containing 0.1% Tween 20, and then incubated overnight at 4Ōäā with the following primary antibodies: antimouse ╬▓-actin, antimouse Bcl-2, antimouse Bax, antimouse PI3K, antirabbit phospho-PI3K (p-PI3K), antimouse Akt, and antimouse phospho-Akt (p-Akt) (1:1,000; Santa Cruz Biotechnology). Subsequently, the membranes were incubated for 1 hour with secondary antibodies (1:2,000; Vector Laboratories), and band detection was performed using the enhanced chemiluminescence detection kit (Santa Cruz Biotechnology). The bands were quantified using an Image-Pro Plus computer-assisted image analysis system (Media Cyberbetics Inc., Silver Spring, MD, USA).
Data Analysis
To compare the relative expression of bands, densitometric analysis was performed using Molecular Analyst ver. 1.4.1 (Bio-Rad). Cell counting and optical density measurements were performed using Image-Pro Plus computer-assisted image analysis system attached to a light microscope (Olympus Co., Tokyo, Japan).
All data were analyzed using IBM SPSS Statistics ver. 21.0 (IBM Co., Armonk, NY, USA), and data are expressed as the mean┬▒standard error of the mean. For comparisons among groups, a one-way analysis of variance and Duncan post hoc test were performed with P<0.05 as an indication of statistical significance.
RESULTS
Effects of Berberine on Short-Term Memory
The results of step-down avoidance task are presented in Fig. 1. Latency was 149.38┬▒10.37 seconds in the sham-operation group, 11.50┬▒3.16 seconds in the ischemia-induction group, 63.13┬▒20.13 seconds in the ischemia-induction and 20 mg/kg berberine-treated group, 61.63┬▒2.16 seconds in the ischemia-induction and 50 mg/kg berberine-treated group, and 114.00┬▒17.71 seconds in the ischemia-induction and 80 mg/kg berberine-treated group. The present results showed that short-term memory was deteriorated with the induction of ischemia (P<0.05), but that treatment with berberine alleviated ischemia-induced short-term memory impairment in the ischemic gerbils (P<0.05).
Effects of Berberine on GFAP and CD11b Expression in the Hippocampal CA1 Region
GFAP and CD11b expression in the hippocampal CA1 region is presented in Fig. 2. When the intensity level of GFAP in the sham-operation group was set at 1.00, GFAP was 1.70┬▒0.05 in the ischemia-induction group, 1.77┬▒0.07 in the ischemia-induction and 20 mg/kg berberine-treated group, 1.27┬▒0.08 in the ischemia-induction and 50 mg/kg berberine-treated group, and 1.11┬▒0.08 in the ischemia-induction and 80 mg/kg berberine-treated group. The present results showed that expression of GFAP in the hippocampal CA1 region was enhanced by the induction of ischemia (P<0.05), but that treatment with berberine reduced GFAP expression in the ischemic gerbils (P<0.05).
When the intensity level of CD11b in the sham-operation group was set at 1.00, the level of CD11b was 2.11┬▒0.09 in the ischemia-induction group, 2.09┬▒0.11 in the ischemia-induction and 20 mg/kg berberine-treated group, 1.52┬▒1.35 in the ischemia-induction and 50 mg/kg berberine-treated group, and 1.35┬▒0.08 seconds in the ischemia-induction and 80 mg/kg berberine-treated group. The present results showed that expression of CD11b in the hippocampal CA1 region was enhanced by the induction of ischemia (P<0.05), but that treatment with berberine reduced CD11b expression in the ischemic gerbils (P<0.05).
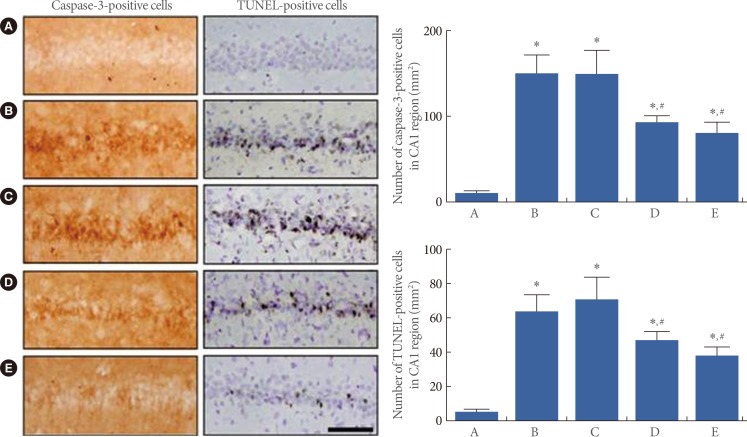
Effects of Berberine on Caspase-3 Expression and DNA Fragmentation in the CA1 Region of the Hippocampus
The numbers of caspase-3-positive and TUNEL positive cells in the hippocampal CA1 region are presented in Fig. 3. The number of caspase-3-positive cells was 7.01┬▒1.85/mm2 in the sham-operation group, 153.15┬▒28.36/mm2 in the ischemia-induction group, 152.06┬▒20.67/mm2 in the ischemia-induction and 20 mg/kg berberine-treated group, 82.50┬▒4.45/mm2 in the ischemia-induction and 50 mg/kg berberine-treated group, and 64.50┬▒10.84/mm2 in the ischemia-induction and 80 mg/kg berberine-treated group. The present results showed that caspase-3 expression in the hippocampal CA1 region was enhanced by the induction of ischemia (P<0.05), but that treatment with berberine reduced caspase-3 expression in the ischemic gerbils (P<0.05).
The number of the TUNEL-positive cells in the CA1 region was 3.01┬▒1.06/mm2 in the sham-operation group, 64.74┬▒8.99/mm2 in the ischemia-induction group, 71.63┬▒11.00/mm2 in the ischemia-induction and 20 mg/kg berberine-treated group, 45.30┬▒3.80/mm2 in the ischemia-induction and 50 mg/kg berberine-treated group, and 34.79┬▒3.63/mm2 in the ischemia-induction and 80 mg/kg berberine-treated group. The present results showed that DNA fragmentation in the hippocampal CA1 region was enhanced by the induction of ischemia (P<0.05), but that treatment with berberine reduced DNA fragmentation in the ischemic gerbils (P<0.05).
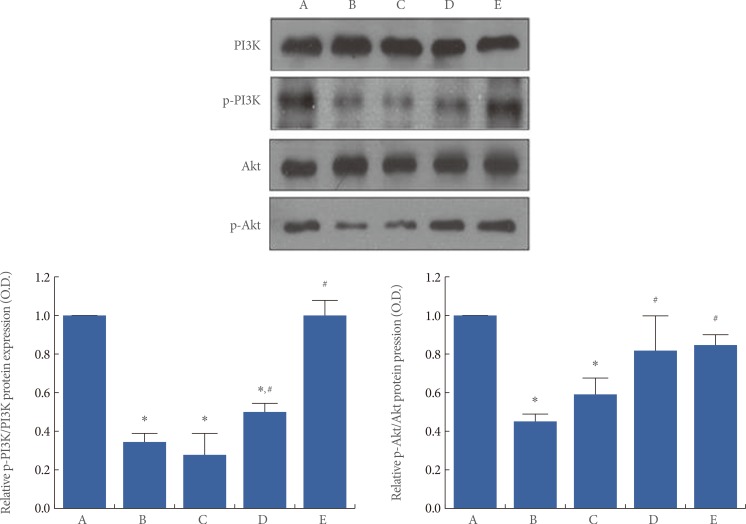
Effects of Berberine on Akt and PI3K Expression in the Hippocampus
We determined the relative expression of PI3K, p-PI3K, Akt, and p-Akt in the hippocampus (Fig. 4). When the ratio of p-PI3K/PI3K (73-82 kDa) in the sham-operation group was set at 1.00, the ratio of p-PI3K/PI3K was 0.34┬▒0.05 in the ischemia-induction group, 0.28┬▒0.10 in the ischemia-induction and 20 mg/kg berberine-treated group, 0.58┬▒0.05 in the ischemia-induction and 50 mg/kg berberine-treated group, and 1.01┬▒0.08 in the ischemia-induction and 80 mg/kg berberine-treated group (Fig. 4). The present results showed that the ratio of p-PI3K/PI3K in the hippocampus was decreased by the induction of ischemia (P<0.05), but that treatment with berberine increased the ratio of p-PI3K/PI3K in the ischemic gerbils (P<0.05).
When the ratio of p-Akt/Akt (60 kDa) in the sham-operation group was set at 1.00, the ratio of p-Akt/Akt was 0.47┬▒0.02 in the ischemia-induction group, 0.59┬▒0.07 in the ischemia-induction and 20 mg/kg berberine-treated group, 0.85┬▒0.10 in the ischemia-induction and 50 mg/kg berberine-treated group, and 0.88┬▒0.03 in the ischemia-induction and 80 mg/kg berberine-treated group (Fig. 4). The present results showed that the ratio of p-Akt/Akt in the hippocampus was decreased by the induction of ischemia (P<0.05), but that treatment with berberine increased the ratio of p-Akt/Akt in the ischemic gerbils (P<0.05).
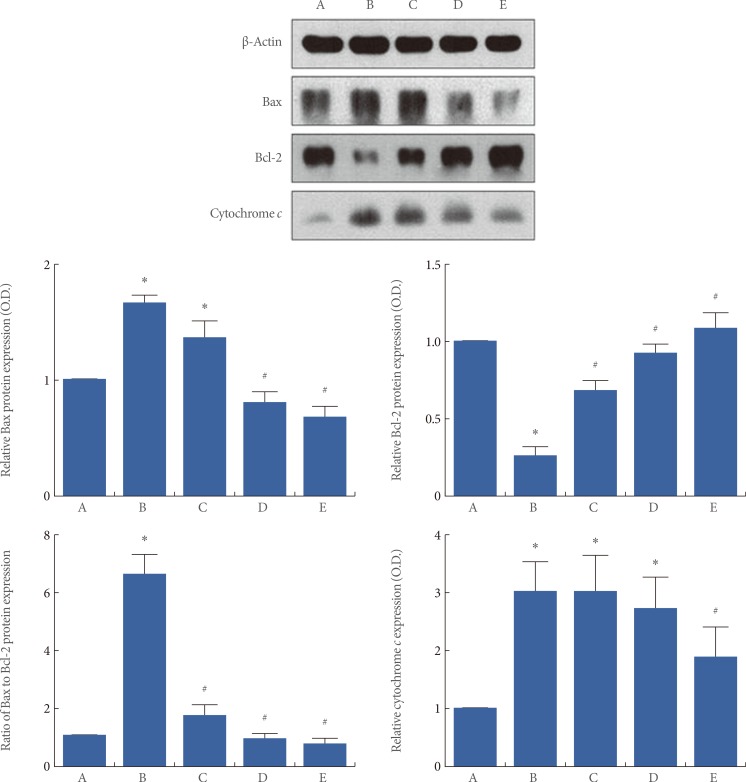
Effects of Berberine on the Expression of Bax, Bcl-2, and Cytochrome c in the Hippocampus
We determined the relative expression of Bax, Bcl-2, and cytochrome c in the hippocampus (Fig. 5). When the level of the Bax (23 kDa) in the sham-operation group was set at 1.00, the level of Bax was 1.65┬▒0.07 in the ischemia-induction group, 1.38┬▒0.15 in the ischemia-induction and 20 mg/kg berberine-treated group, 0.75┬▒0.07 in the ischemia-induction and 50 mg/kg berberine-treated group, and 0.61┬▒0.08 in the ischemia-induction and 80 mg/kg berberine-treated group. The present results showed that the level of Bax expression in the hippocampus was increased by the induction of ischemia (P<0.05), but that treatment with berberine decreased the level of Bax expression in the ischemic gerbils (P<0.05).
When the level of Bcl-2 (26 kDa) in the sham-operation group was set at 1.00, the level of Bcl-2 was 0.28┬▒0.02 in the ischemia-induction group, 0.70┬▒0.02 in the ischemia-induction and 20 mg/kg berberine-treated group, 0.93┬▒0.05 in the ischemia-induction and 50 mg/kg berberine-treated group, and 1.09┬▒0.09 in the ischemia-induction and 80 mg/kg berberine-treated group. The present results showed that the level of Bcl-2 expression in the hippocampus was decreased by the induction of ischemia (P<0.05), but that treatment with berberine recovered the level of Bcl-2 expression in the ischemic gerbils near to the control level (P<0.05).
The ratio of Bax to Bcl-2 was calculated. When the ratio of Bax to Bcl-2 in the sham-operation group was set at 1.00, the ratio of Bax to Bcl-2 was 6.38┬▒0.74 in the ischemia-induction group, 1.74┬▒0.28 in the ischemia-induction and 20 mg/kg berberine-treated group, 0.66┬▒0.11 in the ischemia-induction and 50 mg/kg berberine-treated group, and 0.55┬▒0.13 in the ischemia-induction and 80 mg/kg berberine-treated group. The present results showed that the ratio of Bax to Bcl-2 in the hippocampus was increased by the induction of ischemia (P<0.05), but that treatment with berberine decreased the ratio of Bax to Bcl-2 in the ischemic gerbils (P<0.05).
When the level of cytochrome c (15 kDa) in the sham-operation group was set at 1.00, the level of cytochrome c was 3.07┬▒0.46 in the ischemia-induction group, 3.16┬▒0.57 in the ischemia-induction and 20 mg/kg berberine-treated group, 2.70┬▒0.44 in the ischemia-induction and 50 mg/kg berberine-treated group, and 1.65┬▒0.24 in the ischemia-induction and 80 mg/kg berberine-treated group. The present results showed that the level of cytochrome c expression in the hippocampus was increased by the induction of ischemia (P<0.05), but that treatment with berberine suppressed the level of cytochrome c expression in the ischemic gerbils (P<0.05).
DISCUSSION
Global cerebral ischemia induces neuronal damage in the hippocampus, and causes deficits in learning ability and memory function [1,11]. Berberine has been shown to protect against ischemic brain injury by decreasing intracellular reactive oxygen species levels and subsequently inhibiting the mitochondrial apoptotic pathway [24]. Berberine also prevents changes in oxidative stress and choline esterase activity, and consequently can improve the memory impairment seen in streptozotocin-induced diabetic rats [16]. In the present study, ischemic insult decreased latency in the step-down avoidance task, indicating that ischemia deteriorated short-term memory. In contrast, berberine enhanced latency in the step-down avoidance task. These results indicate that berberine alleviated the ischemia-induced short-term memory impairment.
Reactive astrogliosis inhibits axonal regeneration and exacerbates apoptotic neuronal cell death following ischemic damage [2]. Additionally, microglia activation is associated with neuronal death in ischemic gerbils [25]. Therefore, it is not surprising that inhibiting microglia activation attenuates injury-induced neuronal death [7,26]. Reactive astrogliosis and microglia activation have been implicated in the neuronal dysfunction and cell death [4,7]. In the present study, ischemia insult increased GFAP and CD11b expression in the hippocampal CA1 region. In contrast, berebrine treatment suppressed ischemia-induced increments of GFAP and CD11b expression, showing that berbeine can attenuate ischemic injury by inhibiting reactive astrogliosis and microglia activation.
Apoptosis has been implicated in the pathogenesis of cerebrovascular disease; specifically TUNEL-positive and caspase-3-positive cells appear to increase in ischemic gerbils and in intracerebral hemorrhagic rats [11,27]. In contrast, decrements of these numbers indicate the suppression of apoptosis [11,27]. In the present study, caspase-3-positive and TUNEL-positive cells in the hippocampal CA1 region were observed to increase following ischemic insult, while berberine suppressed these numbers.
Bcl-2 is also involved in the inhibition of apoptosis, while Bax is associated with apoptosis promotion [28]. Decrements of Bcl-2 expression have been observed in alcohol-induced apoptotic neuronal death in the hippocampus of gerbils [21]. Moreover, enhanced Bax and suppressed Bcl-2 expression represents apoptotic neuronal death in cerebrovascular disease [27]. In the present study, ischemic insult was found to increase Bax expression and decrease Bcl-2 expression, resulting in an increased Bax to Bcl-2 ratio. We also found that treatment with berberine suppressed Bax and enhanced Bcl-2 expression in ischemic gerbils.
The pro- and antiapoptotic members of the Bcl-2 family control the release of cytochrome c. A decreased Bax to Bcl-2 ratio can inhibit cytochrome c release, and consequently suppresses apoptosis [29], since the release of cytochrome c from the mitochondria is known to activate apoptosis. In the present study, while cytochrome c expression was increased in the hippocampus of ischemic gerbils, berberine was observed to decrease this expression.
Based on these results, we found that berberine exerted inhibitory effects against ischemia-induced apoptosis in gerbils through altering apoptosis-related proteins. Wang et al. [18] reported that berberine blocks K+ channels of hippocampal CA1 neurons, which subsequently restores cation balance in neurons following anoxic/ischemic injury. This situation then leads to the suppression of apoptosis and a substantial increase in cell survival. Thus, berberine protects against ischemic brain injury by decreasing intracellular reactive oxygen species and subsequently inhibiting the mitochondrial apoptotic pathway [24].
Akt is a serine/threonine kinase and a major downstream target of PI3K. Activation of the PI3K/Akt pathway can promote angiogenesis and diminish microglial/astrocytic proliferation, resulting in the reduction of infarct volumes and behavioral recovery in ischemic rats [30]. Koh et al. [31] reported that global ischemia decreased Akt phosphorylation in the hippocampal CA1 region, while in contrast, Akt activation exerted a protective effect on global ischemic injury. Neuroprotective and postconditioning effects of transient middle cerebral artery occlusion in rats have also been reported to be maintained through activation of the PI3K/Akt pathway [32]. Furthermore, PI3K/Akt pathway can prompt survival signaling in neonatal rats subjected to hypoxic/ischemic conditions [33]. In the present study, ischemic insult suppressed p-Akt and p-PI3K expressions in the hippocampus. Treatment with berberine enhanced p-Akt and p-PI3K expression in ischemic gerbils, showing that berberine activated the PI3K/Akt pathway. These data suggest that phosphorylation of PI3K and Akt mediate a berberine-induced neuroprotective effect through suppressing apoptosis, and that this contributes to hippocampal cell survival.
In the urological aspect, berberine has been suggested as a novel chemotherapy drug to treat bladder cancer via suppressing tumor growth [34]. Yu et al. [35] demonstrated that berberine pretreatment serves a protective role against hypoxia/reoxygenation induced apoptosis in human renal proximal tubular cells by suppression of mitochondrial and endoplasmic reticulum stress pathways. Taken together, berberine exerts its neuroprotective effect against ischemic insult by inhibiting neuronal apoptosis via activation of the PI3K/Akt signaling pathway. This antiapoptotic effect is achieved through inhibition of reactive astrogliosis and microglia activation. Based on the present results, we suggest that berberine may serve as a therapeutic agent for stroke-induced neurourological problems.