ŌĆó HIGHLIGHTS
- Naftopidil reproducibly induces outward current of IPSC in a subset of substantia gelatinosa neurons.
- The naftopidil response is mediated by glycinergic and/or GABAergic neural transmission.
- Other ╬▒1-adrenoceptor antagonists, particularly prazosin, yield weaker changes in the outward current.
INTRODUCTION
In elderly men, lower urinary tract symptoms (LUTS) are related to benign prostatic obstruction/benign prostatic hyperplasia (BPH), bladder dysfunction including overactive bladder, and nocturnal polyuria [1]. Although LUTS/BPH typically is not a life-threatening condition, the impact of LUTS/BPH on the quality of life can be significant and should not be underestimated [2]. To manage male LUTS pharmacologically, ╬▒1- adrenoceptor blockers are well-established drugs and are prescribed widely. ╬▒1-Adrenoceptor antagonists relieve the voiding symptoms of LUTS by relaxing smooth muscle in the urethra and prostate, resulting in reduction of enhanced tonus or contractility, thus counteracting the lowered urine flow rates [3]. Alpha1-adrenoceptors also participate in improving the storage symptoms of LUTS [4], although the mechanism of action of the compounds in this context remains unclear. In rats subjected to conscious cystometry, tamsulosin, naftopidil, and silodosin all have been shown to prolong the micturition interval [5]. Clinical studies comparing the effects of ╬▒1-adrenoceptor antagonists, naftopidil, and tamsulosin yielded contradictory results in equivalence for the International Prostate Symptom Score [6], but yielded consistent differences for the volume at first desire to void and at maximum desire to void in the urodynamic study [7].
The micturition reflex is believed to be mediated by neuronal circuits in the lumbosacral spinal cord [8-10]. It has been suggested that ╬▒1-adrenoceptor antagonists suppress the micturition reflex via action in the spinal cord by isovolumetric cystometry [11], employing a mode of action of naftopidil that requires functional upregulation of ╬│-aminobutyric acid (GABA) and/or glycine receptors at the lumbosacral level [12]. By using voltage-clamp recordings, it is possible to identify primary afferent information in substantia gelatinosa (SG, lamina II of Rexed) neurons [13] mediated by different primary afferent fibers [14,15]. The efficacy of synaptic transmission is determined by presynaptic neurotransmitter release probability and postsynaptic responsiveness, which are evaluated as the frequency and amplitude of miniature excitatory or inhibitory postsynaptic current (mEPSC or mIPSC), respectively. Patchclamp recording of spinal slices has been used to measure mEPSC and mIPSC, revealing that presynaptic firing corresponds to postsynaptic depolarization and hyperpolarization, respectively. Analyses of mIPSCs have provided extensive information of experimental interest, as follows. Increments in the frequencies of mIPSCs mean that stimuli act on presynaptic sites, leading to the release of inhibitory neurotransmitters. Increments in the amplitudes of mIPSCs mean that stimuli act on postsynaptic sites, leading to upregulation of the sensitivity of the action sites to inhibitory neurotransmitters. Changes of membrane current indicate outward or inward orientation of the current (hyperpolarization or depolarization) regarding inhibitory neuronal transmissions [16]. Generally, outward IPSCs indicate an influx of Cl-, which induces hyperpolarization.
A previous study facilitated our understanding of the effect of naftopidil on the micturition reflex [17], indicating that naftopidil increases the frequency of mIPSCs by activating inhibitory interneuron terminals in the spinal dorsal horn [17]. However, information on outward/inward currents in IPSCs and differences induced by various ╬▒1-adrenoceptor antagonists (e.g., naftopidil, tamsulosin, and silodosin) remain unclear, naftopidil was the first ╬▒1-adrenoceptor antagonist evaluated in investigation of the mechanism of micturition reflex in the lumbosacral cord. Our primary goal was to determine the effects of an ╬▒1-adrenoceptor antagonist, naftopidil, on outward/inward currents in IPSCs. The secondary goals were to determine the differences between the outward currents induced by naftopidil and the other ╬▒1-adrenoceptor antagonists, and to determine whether the outward current induced by naftopidil mediates GABA/glycine receptor activation in vitro in SG neurons derived from rat lumbosacral spinal cord.
MATERIALS AND METHODS
Spinal Cord Slice Preparation
The methods for obtaining slices of the adult rat spinal cord and for blind patch-clamp recordings from SG neurons have been described in detail elsewhere [17-20]. Briefly, adult male Sprague-Dawley rats (6ŌĆō8 weeks old) were deeply anesthetized with urethane (1.2 g/kg, intraperitoneally), and a lumbosacral laminectomy then was performed. The lumbosacral segments of the spinal cord (L2ŌĆōS3) with ventral and dorsal roots were removed and placed in ice-cold Krebs solution equilibrated with 95% O2ŌĆō5% CO2. The Krebs solution contained (in mM): NaCl 117, KCl 3.6, CaCl2 2.5, MgCl2 1.2, NaH2PO4 1.2, NaHCO3 25, and glucose 11 (pH, 7.4). Immediately after removal of the spinal cord, the rats were killed by exsanguination under urethane anesthesia. The pia-arachnoid membrane was removed after cutting all the ventral and dorsal roots. The spinal cord was mounted on a vibratome, and a 500-┬Ąm-thick transverse slice with the attached dorsal root was cut. The slice was placed on a nylon mesh in the recording chamber in a volume of 0.5-mL Krebs solution, and was completely submerged and perfused with Krebs solution saturated with 95% O2ŌĆō5% CO2 at 37┬▒1┬░C and a flow rate of 10ŌĆō15 mL/min.
Whole-Cell Patch-Clamp Recordings From SG Neurons
The SG was easily discernible with transmitted illumination as a relatively translucent band across the dorsal horn in the transverse slice preparations. Blind whole-cell voltage-clamp recordings were made from SG neurons, as described previously [17,18,20,21]. The patch pipettes were filled with a solution containing cesium solution (in mM): Cs2SO4 110, tetraethylammonium (TEA) 5, CaCl2 0.5, MgCl2 2, ethylene glycol tetraacetic acid 5, 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid 5, and adenosine 5'-triphosphate-Mg 5 (pH, 7.2). With the cesium pipette solution, a holding potential was easily depolarized to 0 mV, since the K+ channels were blocked by cesium and TEA. The tip resistance of the patch pipettes was 6ŌĆō12 M╬®. Series resistance was assessed according to the response to a 5-mV hyperpolarizing step. This value was monitored during the recording session, and data were rejected if values changed by >15%. Signals were acquired with a patch-clamp amplifier (Axopatch 700A, Molecular Devices, Union City, CA, USA). The data were digitized with an AD/DA converter (Digidata 1321A, Molecular Devices), stored on a personal computer using a data acquisition program (Clampex, version 9.0, Molecular Devices), and analyzed using a software package (Clampfit, version 9.0, Molecular Devices). Cell recordings were made in voltage-clamp mode at holding potentials of 0 mV to record IPSCs [17,18,20]. We used a total of 17 animals in this study. One slice was collected from each rat and recordings were obtained for 1ŌĆō5 cells/slice. One to 5 cells were recorded for each set of experiments.
Drug Application
Naftopidil (final concentration, 100╬╝M) (PubChem CID: 4418) (Asahi Kasei Pharma Co., Tokyo, Japan) was dissolved in 1% dimethyl sulfoxide (PubChem CID: 679) (Wako, Osaka, Japan) in Krebs solution. Tamsulosin (100╬╝M) and silodosin (30╬╝M) were dissolved in Krebs solution. The other compounds used in this study were prazosin (10╬╝M) (PubChem CID: 4893) (Sigma-Aldrich, St. Louis, Mom USA), tetrodotoxin (TTX) (1╬╝M) (PubChem CID: 6324668) (Wako), strychnine (2╬╝M) (PubChem CID: 441071) (Sigma-Aldrich), and bicuculline (10╬╝M) (PubChem CID: 10237) (Sigma-Aldrich). All drugs were applied by sequential perfusion in individual cells with washout periods via a three-way stopcock without changes in the perfusion rate or temperature. The application schedule was described previously [22].
Statistical Analysis
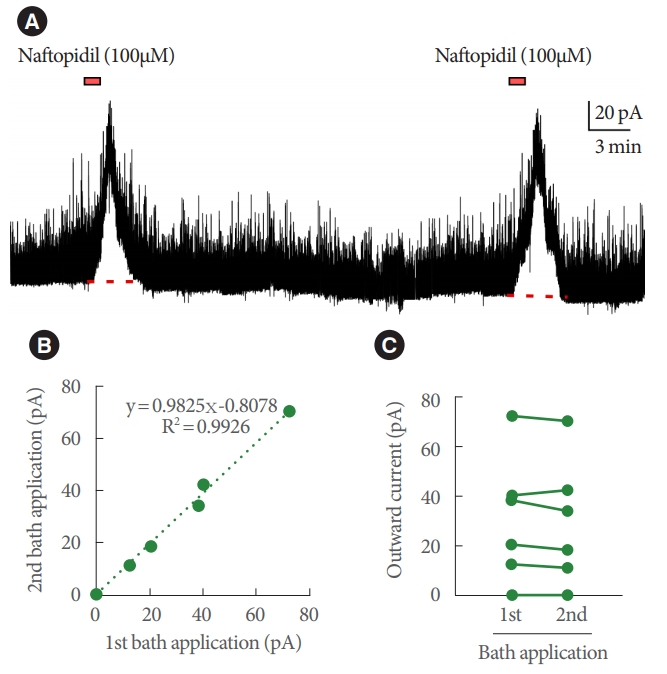
Statistical analysis was performed using JMP ver. 14 (SAS Institute, Cary, NC, USA). Continuous values are revealed mean┬▒ standard error of the mean. Inferential statistical analysis was not performed, because the number of SG neurons that exhibited induced outward current was too small. Regarding reproducibility for the outward current induced by naftopidil, correlations were subjected to linear regression analysis, and the values of r2 and the slopes of regression lines were calculated between the 1st and 2nd bath applications in a given neuron. Pearson correlation coefficients (r-values) were analyzed.
RESULTS
Outward/Inward Current of IPSC in SG Neurons Following Bath Application of Naftopidil
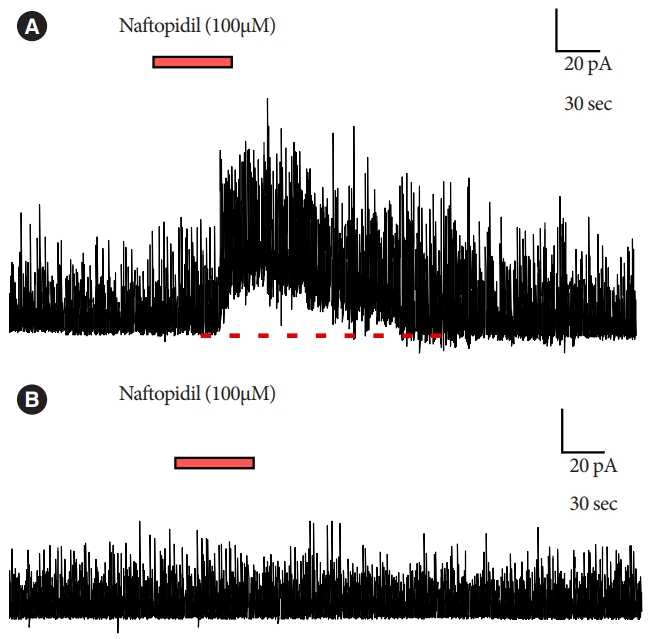
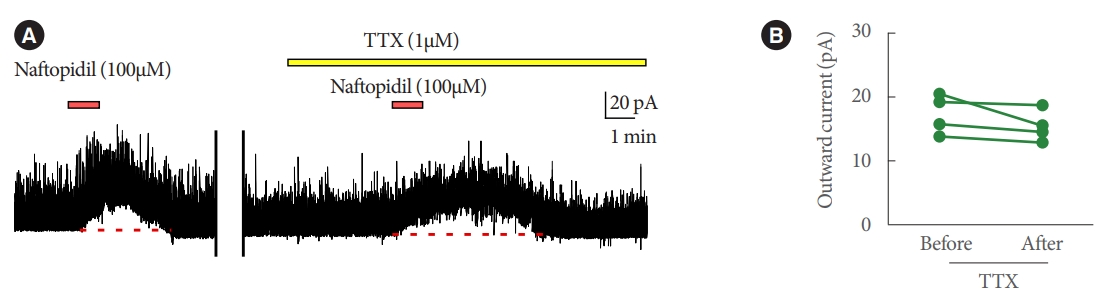
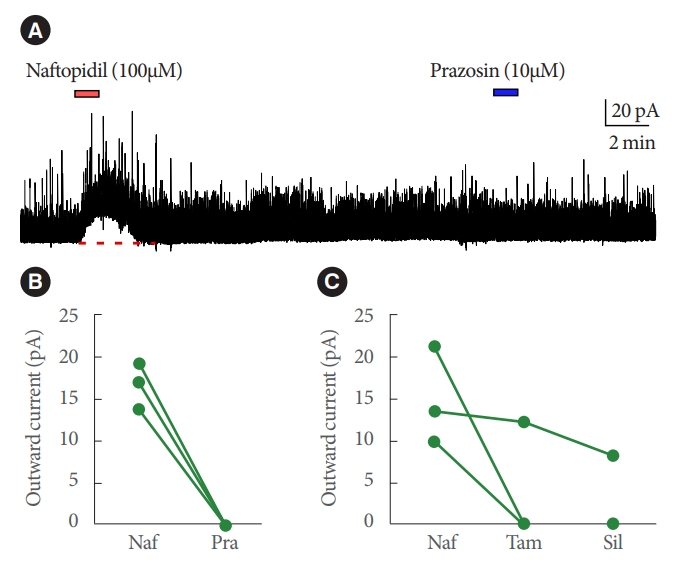
Bath application of naftopidil-induced outward current of IPSC in 13 of 52 SG neurons; typical charts are shown in Fig. 1. The observation of the current was reproducible (Fig. 2). Regression analysis of the currents yielded a Pearson correlation coefficient of 0.996 when comparing between the 1st and 2nd applications of naftopidil; the data plotted as a line with a slope of 0.983. The naftopidil-induced outward current also was observed in the presence of TTX (Fig. 3). Inward current of IPSC was not observed in any SG neurons following naftopidil exposure.
Impact of Glycinergic and GABAergic System for GABA/Glycine-Mediated Outward Current of IPSC Generated by Naftopidil in SG Neurons
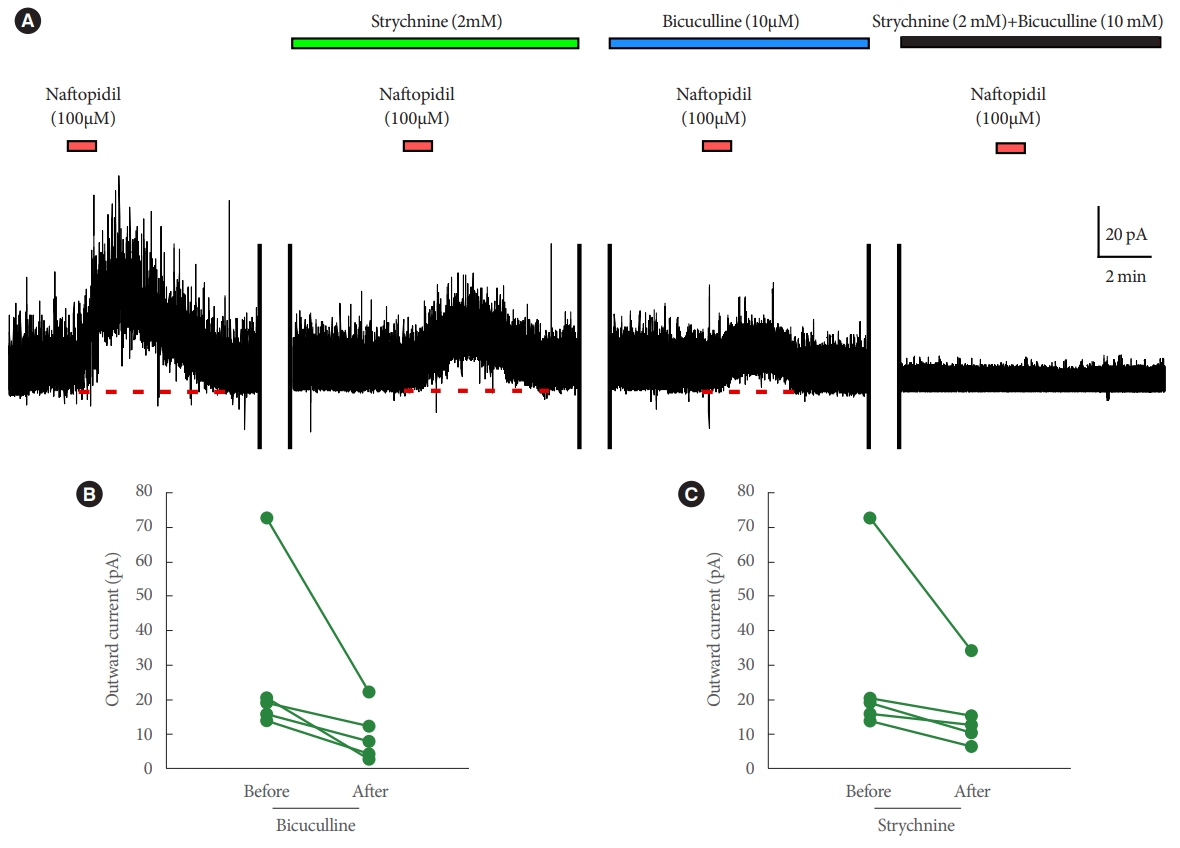
The outward current induced by naftopidil in each SG neuron was attenuated in the presence of strychnine (from 28.4 pA┬▒5.0 pA to 15.8 pA┬▒2.2 pA, n=5) or bicuculline (from 28.4 pA┬▒5.0 pA to 9.8 pA┬▒1.6 pA, n=5) (Fig. 4). When both strychnine and bicuculline were applied simultaneously, no naftopidil-induced outward current was detected (n=5) (Fig. 4).
Effects of Other ╬▒1-Adrenoceptor Antagonists on the GABA/Glycine-Mediated Outward Current of IPSC
Next, the effects of other ╬▒1-adrenoceptor antagonists were tested (separately) in 6 SG neurons that exhibited naftopidil-induced GABA/glycine-mediated outward current. In a subset of 3 naftopidil-responsive SG neurons, only 1 of the 3 exhibited outward current in response to tamsulosin or silodosin; notably, the same neuron responded to both compounds. In the remaining 3 naftopidil-responsive SG neurons, prazosin did not induce an outward current (Fig. 5).
DISCUSSION
In the present study, only a subset of SG neurons was found to be naftopidil responsive. The outward current induced by naftopidil was observed reproducibly and was not antagonized by TTX (the primary endpoint). The outward current was suppressed by strychnine and/or bicuculline, and the current was evoked at lower probabilities by tamsulosin and silodosin than by naftopidil (the secondary endpoints). Prazosin did not induce an outward current in SG neurons know to be naftopidil responsive. The inward current was not observed by naftopidil.
The Effects of Naftopidil on the Outward Current in IPSC and the GABA/Glycine-Mediated Effects in SG Neurons
Regarding with the primary endpoint, naftopidil-induced outward current of IPSCs in a subset of SG neurons. It has been speculated that this variability in neuron responsivity reflects classification of SG neurons into (at least) 4 types [20], such that only certain SG neurons are naftopidil responsive. We infer that naftopidil secondarily suppresses postsynaptic neuronal activity. Since the ability of naftopidil to induce the outward current likely was not altered in the presence of TTX (Fig. 3B), the current presumably is not associated with sodium channels in the synapse.
Regarding with the secondary endpoint, the outward current induced by naftopidil was attenuated by pre-exposure to strychnine and/or bicuculline before naftopidil. These data suggest that naftopidil induces the GABA/glycine-mediated outward current (i.e., hyperpolarization) via glycinergic and/or GABAergic neurotransmission [17]. This interpretation is supported by a previous report that showed that strychnine and bicuculline eliminated the frequency of mIPSCs that was induced by naftopidil [17]. As shown in Fig. 1, mild facilitation of frequency was observed for IPSCs together with outward current in the SG neuron; this result indicated that induction of the SG neurons likely is suppressed by naftopidil, perhaps via enhancement of presynaptic release of GABA and/or glycine in addition to hyperpolarization. Therefore, naftopidil may suppress neural activity multimodally. A subpopulation of naftopidil-responsive SG neurons did not exhibit GABA/glycine-mediated outward currents of IPSCs upon exposure to tamsulosin, silodosin, or (notably) prazosin. The basis of this distinction in antagonist sensitivity is unclear, but it is possible that one or more ╬▒1- adrenoceptor antagonists, or conceivably the ╬▒1-adrenoceptor itself, are excluded from the site of action for induction of the GABA/glycine-mediated outward current of IPSCs. Naftopidil may attenuate bladder overactivity by enhancing fine control of excess or abnormal urge signals in the spinal cord, altering hyperpolarization in SG neurons.
The Role of GABA/Glycine-Mediated Outward Current of IPSC
IPSC is induced by inhibitory neurotransmitters (e.g., glycine and GABA) that are released from presynaptic terminals, resulting in an influx of ClŌĆō into the postsynaptic neuronal cells. Decreasing of the membrane potential of the postsynaptic cells by ClŌĆō influx generates an outward current. Simultaneously, hyperpolarization is observed, and the responses of postsynaptic cells are suppressed. In rat, the infusion of 0.1% acetic acid into the urinary bladder induces a nociceptive transmission, as revealed by upregulation of c-Fos expression at the L6 level of the spinal cord [23]. Acetic acid also induces shortening of the inter contraction interval in rats subjected to conscious cystometry [24]. Those investigations indicated that interruption of supraspinal modulation enhances nociceptive transmission in the lower urinary tract [23]. A portion of the superficial neurons receives both nociceptive and nonnociceptive inputs from the lower urinary tract, as shown by upregulation of c-Fos expression [25]. Those results indicated the possibility that a primary afferent signal is transmitted from the bladder epithelium to the superficial SG neurons in the spinal cord. Therefore, outward currents and IPSCs in the SG neurons can be considered reasonable targets for exploring the pathology and treatment of LUTS.
Extrapolation of Induction of GABA/Glycine-Mediated Outward Current of IPSC to Micturition
GABA/glycine-mediated outward current of IPSC, which reflects hyperpolarization, is one mechanism for suppressing signal transmission by neuronal cells. The nociceptive stimuli are suppressed by hyperpolarization with resulting decreases in the excitability of SG neurons [26]. These effects could suppress the micturition reflex at the spinal cord. In the context of LUTS with pain (e.g., painful bowel syndrome), the importance of GABA/glycine-mediated outward current of IPSC will need to be further investigated. These concepts may clarify the mechanism of pathogenesis and evaluation of treatments for these symptoms.
In conclusion, a subpopulation of rat SG neurons, prepared from the L6ŌĆōS1 level of the spinal cord, were shown to exhibit GABA/glycine-mediated outward current of IPSC in response to naftopidil exposure. This result suggested that ╬▒1-adrenoceptor antagonists, particularly naftopidil, may induce neural suppression, in part by mediating hyperpolarization. The observed response was associated with glycinergic and/or GABAergic neural transmission, but may not depend on ╬▒1-adrenoceptors themselves. The present findings suggest that naftopidil may alleviate storage symptoms while also providing hyperpolarization in SG neurons, suppressing the micturition reflex by acting at the level of the spinal cord.
This study has some limitations. Although afferent nerves from the urothelium largely project to lamina X in the dorsal horn, SG neurons in lamina II were used. As described previously, a portion of the superficial neurons receives both nociceptive and nonnociceptive inputs from the lower urinary tract, as evidenced by upregulation of c-Fos expression [25]. Although the use of lamina X would be ideal, that structure has a low density of neurons, making the blind attachment of patchclamp electrodes more difficult. Instead, lamina II was examined.